ДИНАМІКА ПОПУЛЯЦІЙ
Спеціалісти в галузі екології вкладають у поняття динаміки популяції різний зміст. Дійсно, це дуже ємне поняття. Динамічний популяційний процес включає в себе кілька відносно самостійних явищ: зміна розмірів території, яку займає популяція, зміна густоти особин у межах популяційного поля, межі при цьому можуть залишатися попередніми, і, нарешті, зміна розмірів запасу фітомаси в розрахунку на одиницю площі внаслідок трансформації стану особин, що входять до популяції.
Якщо все ж таки динаміку популяцій розуміти тільки як зміну чисельності особин, то трансформація властивостей особин, зміщення співвідношення між різними їхніми групами і особливо функціонування популяції будуть тоді виступати як механізми, що забезпечують динаміку чисельності. Коливання чисельності особин у популяціях, що є типовими для багатьох рослин та тварин, називаються флуктуаціями.Загальний хід зміни чисельності особин у популяції визначається
рівнянням:
де N - чисельність особин у популяції, В - народжуваність, D - смертність, / - імміграція, Е - еміграція, t - час.
Розміри популяції можуть зростати або за рахунок високої народжуваності, або за рахунок високої імміграції, або за рахунок поєднання обох цих факторів. Знижує розмір популяції смертність та еміграція особин за її межі.
Народжуваність - це кількість нових особин, які з’являються в популяції за певний період часу в розрахунку на одну особину. Для порівняльної характеристики народжуваності використовують особливий показник - питому народжуваність, яку обчислюють як кількість новонароджених особин, що віднесена
до початкового розміру популяції. Величина народжуваності суттєво змінюється в різних біологічних видах. Наприклад, в оселедця одна особина відкладає до 50 тис. ікринок, а в акул -тільки декілька штук.
Звичайно народжуваність пов’язана з наявністю турботи про потомство. Якщо така турбота є, то народжуваність нижча. Широко варіює цей показник у рослин. Орхідні продукують до мільйона насінин, але вони пилоподібні й не мають добре диференційованого зародку. А в деяких видів пальм за рік утворюється всього кілька плодів, але кожний із них важить більше кілограма, вони мають великий зародок та значний запас поживних речовин. Природний добір спрямований не на звичайне збільшення плодовитості, а на таке співвідношення плодовитості з характером зачатків розмноження, яке забезпечує найбільшу життєздатність потомства.Рівень народжуваності помітно змінюється протягом років. Добре відомі цикли плодоношення яблунь. У дрібних гризунів (лемінги, миші) спалахи високої народжуваності відбуваються один раз на 3-4 роки, їм відповідають і спалахи народжуваності в популяціях хижаків - лисиць, песців, сов. У сарни піки народжуваності спостерігаються в середньому один раз на 40 років.
Смертність особин у популяціях ще недостатньо вивчена. Одні фахівці гадають, що старіння і смерть особин генетично запрограмовані та мало залежать від умов існування організмів. Інші вчені вважають, що будь-який організм потенційно безсмертний. Старіння та смерть є наслідком накопичення метаболітів, псування мембран та інших процесів, які мають обов’язковий характер. Однак у кожного організму є свої межі біологічного життя. Вимирання особин у популяціях може спостерігатися на різних фазах розвитку особини. Закономірним є вимирання найбільш старих, але пік смертності припадає на молоді особини. Майже у всіх рослин найбільша кількість смертей припадає на фазу проростання насіння, у тварин - на фазу молодняку.
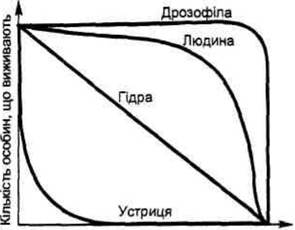
За величиною смертності особин у різні періоди їхнього життя будують так звані криві виживаності, що показують, на який період припадає найбільша смертність особин. Виділяють три основні типи кривих виживаності (рис. 6.5). Перший тип кривих виживаності
споете-
рІГЯЄТЬСЯ у рЯЗІ ОДНЭКОВОГО рОЗ- рис 65 різні типи кривих
міру смертнОСті в будь-якОму виживанОСті
віці організму, він описується монотонно падаючою кривою і спостерігається в популяцій, що живуть у сприятливих умовах (наприклад, гідра). Другий тип кривих виживаності відрізняється високою смертністю молодих особин на перших фазах їхнього життя.
Але особи, що вижили, вже існують до природної смерті. Цей тип кривих виживаності характерний для більшості рослин і тварин (наприклад, устриця). Третій тип кривих виживаності властивий організмам, в яких пік смертності припадає на старі особини. Такі криві властиві деяким комахам (наприклад, дрозофілам), спостерігається він і в популяціях людини.Звичайно, реальний тип виживаності того чи іншого виду організмів не зводиться до одного з основних типів - виживаність для кожного організму є суворо індивідуальною, як це можна
бачити з кривих вижива-
25 50 75 100% НОСТІ, Наведвно
на рис. 6.6.
Динаміка зміни чисельності особин в популяціях може мати різноманітний характер. Якийсь час ріст популяції може бути екс-поненційним, але більш реалістичні моделі сигмоїд- ного росту або j-подібного. Ріст популяцій відповідно до сигмоїдних кривих найчастіше спостерігається в популяціях бактерій та дріжджів. Він може бути описаний рівнянням: 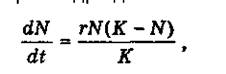
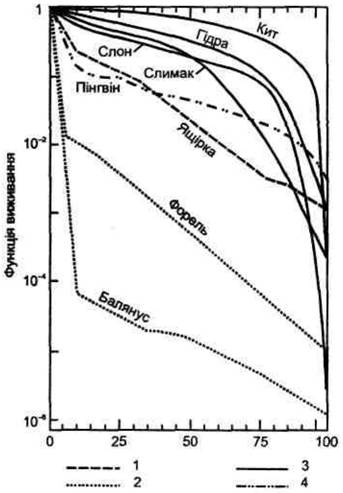
Рис. 6.6. Основні типи кривих виживання організмів різних видів:
1 - постійна смертність у будь-якому віці: 2 - найбільша смертність на початку життя: 3 - найбільша смертність у кінці життя:
4 - переважаюча смертність в перші і останні роки життя (за даними В.П. Кучерявого, 2000)
де dN/dt - швидкість зміни чисельності особин у популяції в часі, г - природжена швидкість росту популяції, що визначається біологічними особливостями виду. Високі значення г вказують на високу швидкість розмноження. К — ємність середовища щодо даної популяції, вона чисельно може бути рівною максимально можливій у даних умовах кількості особин у популяції.
Стан популяції при її сигмоїдному рості залежить від співвідношення параметрів N та К. Якщо N*> К, швидкість росту популяції стає від'ємною величиною і кількість особин у ній зменшується. При N < К швидкість росту популяції додатна і кількість особин у ній прогресивно зростає.
При N = К розміри популяції стабільні.
зростає експоненційно, а при від'ємному значенні - кількість особин у ній зменшується. Ріст популяцій за /-подібною кривою іноді називають ростом типу «бум/крах». Дійсно, у цьому разі зростання кількості особин у популяції
162
відбувається швидко, але потім у момент виснаження ресурсів починається різкий спад. У цього типу росту немає положення стабілізації чисельності особин у популяції.
Є два основні варіанти росту популяції: незалежний та залежний від густоти. У першому варіанті кількість особин у популяції зростає незалежно від густоти популяції, у другому -відтворення має зворотний зв'язок: чим більша густота популяції, тим нижча народжуваність. За певної порогової величини показника густоти народжуваність дорівнює смертності, і кількість особин у такій популяції стабілізується.
Задля стійкого існування популяції оптимальною є певна специфічна для даного біологічного виду і типу середовища густота. При розрідженому заселенні популяційного поля ускладнюються зустрічі партнерів по розмноженню, територію важко оберігати від вторгнення конкуруючих особин. Перенаселення, навпаки, створює дефіцит поживних речовин, а у тварин викликає стреси та досить часті сутички між особинами. Перенаселення настільки несприятливе, що в процесі еволюції виробилося багато різних механізмів його запобігання.
6.10.