НЕКОТОРЫЕ ПОКАЗАТЕЛИ СТРУКТУРНОЙ ОРГАНИЗАЦИИ ЖИВОТНОГО НАСЕЛЕНИЯ ПОЧВ
Рассмотренные показатели отвечают лишь задачам сугубо сравнительного анализа в пределах рассматриваемой совокупности выделов. Необходимы также методы, позволяющие рассчитывать соотношения, дифференцированность компонентов каждой группировки, характер распределения величин обилия по видам.
Известная формула биотического разнообразия Фишера (Fisher et al., 1943; Рудяков, 1964; Федоров, 1970) в этом плане может иметь лишь ограниченное применение, так как в ней не учитывается характер количественной дифференцированности компонентов, а лишь общая численность и количество видов. В последнее время в гидробиологии стали применяться информационные индексы разнообразия (Margalef, 1968, и др.). Можно пользоваться формулой
где N— величина суммарного обилия для всей группировки (количество особей, масса), Na, Nb... Ne— частные величины обилия отдельных компонентов, D — разнообразие сообщества в битах на индивид или другую единицу обилия, например, массы. D тем больше, чем большее число видов (или компонентов иного ранга) в исследуемой группировке имеет высокое обилие, т. е. в полидо- минантных сообществах. D будет стремиться к нулю при приближении N к Na, т. е. с увеличением степени монодоминантности. Безусловно, величину численности в этой формуле можно заменять любым другим показателем обилия или энергетических потенций—массой, интенсивностью метаболизма, что абсолютно необходимо при анализе разноразмерных совокупностей видов. 6 нашей литературе уже опубликованы результаты расчетов информационных индексов разнообразия по величинам массы (Гиляров, 1969; Кожова, 1969; Ананьева, 1971).
Информационные индексы разнообразия весьма показательны при сравнительных анализах серий группировок, отражающих последовательные сукцессионные стадии, или ряды по профилю ландшафта, в которых наблюдаются существенные изменения характера дифференцированности компонентов и синтетических величин обилия.
А. М. Гиляров (1969, 1969а) установил отрицательную корреляцию между суммарной биомассой и разнообра208
зием для группировок гидробионтов. Логично предположить, что это соотношение будет наблюдаться лишь в пределах относительного оптимума, а в специфических условиях при низком уровне массы величина индекса D будет уменьшаться. При анализе распределения коллембол в связи с динамикой растительности в пятнистой тундре С. И. Ананьева (1971) получила максимальные величины индекса на промежуточных этапах зарастания пятен при средних величинах суммарной массы. Это Согласуется и с максимальным экологическим разнообразием коллембол на этой стадии и с наиболее смешанным характером растительного покрова (рис. 24). Такая же тенденция отражена и на рис. 25, где представлены данные для ландшафтной серии группировок. Минимальные значения индекса D получены для наиболее специфичных по гидротермическим режимам группировок — наиболее увлажненных и наиболее сухих — под пологом дубравы, в днищах логов и на пастбище, при максимальном и минимальном уровнях общей зоомассы. Исходя из этих данных, можно сделать вывод, что максимальные величины индекса тяготеют к группировкам, формирующимся в условиях, наиболее «усредненных», при максимальной смешанности разнородных факторов. Именно таким характеристикам соответствуют средние стадии зарастания пятен, поляны, луговая степь. В лесостепи максимальная величина индекса получена именно для полян с относительно высокой зоомассой, но, очевидно, с наиболее смешанным составом растительности и животного населения (луго-лесо-степного характера). При суммарных количественных характеристиках животного населения практически невозможно получать индексы с учетом полного видового состава всех групп, во-первых, из-за колоссального объема расчетов, во-вторых, вследствие неизбежной неравномерности систематической обработки различных групп наземных беспозвоночных. В этих случаях можно ограничиться данными по семействам, родам или экологическим группам.
Нам представляется не совсем удачным название этого показателя в применении к сообществам. Правильнее было бы его называть «индексом дифференцированности». Не следует также переоценивать его показательность при сравнительных анализах. Так, два сообщества могут иметь близкие коэффициенты при существенных различиях уровня обилия и видового богатства. В других случаях группировка с высокой дифференцированностью по значительному числу видов может иметь небольшой индекс вследствие преобладания одного из них, тогда как в выделе с низким суммарным обилием при небольшом количестве видов, но в силу отсутствия заметного резкого преобладания одного из них величина коэффициента может быть большей.Интересные данные, демонстрирующие особенности внутренней структурной организованности животного населения, можно получить, используя принцип построения диаграммы Раункиера
209

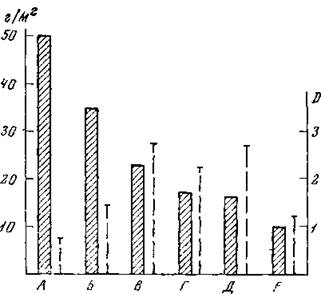
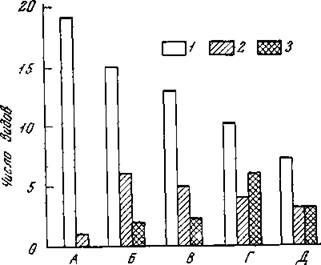
Рис. 24. Информацйоййые индексы разнообразия всей совокупности беспозвоночных, учтенных в почвенных пробах (я), и группировок коллембол (б) в подзоне типичных тундр Западного Таймыра (Чернов, 1973)'
А — пятна голого грунта пятнистой тундры; Б — зарастающие пятна; В — осоково-Моховая бугорковая тундра, /' — разнотравно кустарничковые группировки- Д — луговые группировки
Рис. 25. Суммарная масса (столбики) и индекс (D) разнообразия (прерывистые линии) группировок беспозвоночных, учтенных методом ручной разборки в лесостепи под Курском (Чернов, 1971)
А — дубрава, Б —динще лога в степи; В — поляна; Г — некоснмая луговая степь; Д — косимая луговая степь; Е— пастбище
Рис. 26. Распределение видов коллембол по классам обилия в лесостепи под Курском (по материалам К). Б. Бызовой)
Л — косимая луговая степь, Б — некосимая луговая степь; В — дубрава; Г—днище лога в степи, Д — овсяное поле 1, 2, 3 — классы обилия по убыванию
(Беклемишев, 1931; Грейг-Смит, 1967).
Причем вместо весьма формального (Беклемишев, 1931) показателя встречаемости можно использовать величины обилия (численность, масса, процентные соотношения и т. д.). Разбив виды (или другие компоненты) по классам обилия и отложив последние на оси абсцисс, а на оси ординат — количество видов и групп, мы получим диаграмму, которая будет отражать тенденции к поли- или монодоминантности, видовую насыщенность и равномерность дифференциации состава, определяющую степень стабильности. При достаточно большом количестве классов, как правило, наблюдается та же тенденция к бимодальному распределению (вершины в минимальном и максимальном классах), что и в кривой Раункиера по встречаемости. Эта тенденция к расслоению на крайние варианты по величинам обилия — одна из характерных закономерностей структуры сообществ, требующая специального изучения. Наибольший подъем левой части (области минимальных классов) при отсутствии максимального соответствует группировкам с наибольшей степенью разнообразия и устойчивости, тенденция к (понижению левой части диаграммы при достаточно высокой правой будет наблюдаться в группировках, формирующихся в условиях, отклоняющихся от оптимума, когда в силу «биоценотического закона» Тинемана (Thienemann, 1925), наряду со снижением видового разнообразия, обычно резко повышается обилие отдельных форм. Примером таких закономерностей могут служить данные по коллем- болам Стрелецкого участка Центрально-Черноземного заповедника (рис. 26). Для большей наглядности и простоты мы выделили всего три класса обилия. И даже при таком грубом подразделении получилась картина, четко соответствующая логическим предположениям; наименьшей амплитудой по величине обилия характеризуется группировка косимой степи, где совсем нет видов, относящихся к третьему классу обилия. Степень доминантности здесь наименьшая, что согласуется и с максимальным флористическим богатством растительности именно на косимых участках (необходимо принять во внимание недоучет редких форм). В противоположность этому на дне лога и овсяном Поле при относительно небольшом видовом богатстве значительное количество видов относится к максимальному классу обилия, что отражает большую специфичность этих местообитаний. Очевидно, тенденция к повышению обилия отдельных видов на фоне относительно бедного видового состава— вообще характерная черта разнообразных специфичных группировок, в особенности антропогенных (Бей-Биенко, 1957). Интересно, что даже при трех классах обилия в некоторых группировках отчетлива тенденция к бимодальяости.При анализе количественной дифференцированности животного населения исследователь постоянно сталкивается с понятием доминирцвания, которое в настоящее время интерпретиру-
ется в очень широком произвольном смысле и требует конкретизации. Целесообразно, наряду с известным индексом доминирования (Беклемишев, 1961), выделить несколько специальных показателей, отражающих характер доминантности, в частности, коэффициент доминантности—отношение суммарного обилия эмпирически выделенных доминантов к общему обилию всех видов. Используя для этого рис. 26, легко видеть, что наиболее специфичные группировки (дно лога и овсяное поле) будут иметь наибольшие величины этого коэффициента.
В заключение коснемся еще одного вопроса, имеющего принципиальное значение в методическом отношении. В настоящее время в исследованиях структуры и типологии сообществ принят сугубо видовой подход, т. е. расчеты всех количественных показателей предполагают, как правило, сравнение по видам. Между тем этот подход явно односторонен. Реальна такая ситуация, когда два сравниваемых однотипных сообщества, например, удаленные варианты лугов, могут не иметь общих видов, в силу чего коэффициенты сходства по обилию видов или по фауне будут иметь нулевые величины. Вместе с тем сопоставление разнотипных сообществ, например леса и луга, может дать высокие величины этих индексов. Легко показать на многих примерах, что вид как таковой по отношению к структуре сообщества— компонент случайный.
Факты смены доминантных видов или доминирования одних и тех же видов в разных сообществах сами по себе говорят лишь о степени их эврибионтности (в разных сообществах вид может занимать различные ниши, иметь различный удельный вес и т. д.), но они мало информативны в отношении структурных особенностей сообщества. Так, известно много случаев доминирования одних и тех же видов в крайне разнотипных сообществах, что, однако, не дает повода говорить о несущественных различиях в их структуре. Все это и побудило нас попытаться использовать различные приемы анализа структуры сообщества не только по видам, но и на уровне любых таксонов. Вместе с тем следует признать, что это лишь первые попытки, требующие дальнейших уточнений. Необходимо как можно шире использовать синтетические количественные характеристики (общий уровень обилия, соотношение экологических групп, характер дифференцированности компонентов и др.). При относительном обилии конкретных количественных Данных по группировкам наземных животных эти-показатели используются пока очень мало. Между тем для характеристики животного населения в ряде случаев такие суммарные данные бывают более репрезентативны, чем детальные списки с полным видовым анализом.
В настоящей статье автор не стремился дать рбзор всех методов синэкологического анализа, а рассмотрел лишь те показатели, которые, по его мнению, представляют наибольший интерес для обработки количественных данных по почвенной фауне.
312
Еще по теме НЕКОТОРЫЕ ПОКАЗАТЕЛИ СТРУКТУРНОЙ ОРГАНИЗАЦИИ ЖИВОТНОГО НАСЕЛЕНИЯ ПОЧВ:
- 10.1.2. Показатели состояния почв, определяемых при контроле загрязнения почв
- Педохимические показатели состояния загрязненных почв
- 8.1. Показатели воспроизводства населения
- Некоторые вопросы структурного изучения текста
- 28. Система показателей доходов населения
- применение показателей ферментативной активности в целях мониторинга и диагностики почв Юга России[2]
- 2.1.1 Пример определения индивидуальных показателей структурных сдвигов
- 2.4.1 Пример определения показателей относительных структурных сдвигов
- 2.3.1 Пример определения показателей абсолютных структурных сдвигов
- 10.1. Виды и причины миграций, показатели территориальной подвижности населения
- 3.2.1 Пример определения показателей структурных различий
- Некоторые соображения об обращении с местным населением восточных областей
- Понятие социальной организации. Характеристики организации.Роль организации в социуме.Типология организаций. Формальные и неформальные организации.Структура организации, ее влияние на поведение индивидов. Функции организации.Иерархии в организации. Управление в организации.«Идеальный тип бюрократии» по М. Веберу.