СРАВНИТЕЛЬНЫЙ АНАЛИЗ ГРУППИРОВОК ПОЧВЕННЫХ БЕСПОЗВОНОЧНЫХ
При сравнительных анализах группировок наземных животных (наряду с простейшими индексами доминирования, встречаемости) в нашей литературе достаточно широко используются лишь коэффициенты фаунистического сходства.
Наиболее часто употребляются попарные коэффициенты Жаккара и Серенсена. Первый из них рассчитывается по формуле:
где С—число видов, общих для двух сравниваемых группировок; А — число видов в первой группировке; В — то же, во второй. Формула Серенсена при тех же обозначениях имеет вид:

Эти коэффициенты можно выражать как в долях, так и в процентах. Разработан ряд приёмов дальнейшей обработки и сопоставления полученных попарных индексов, о чём подробно будет сказано ниже, в разделе об аналогичных попарных индексах при количественных анализах. Оба индекса широко используются зоологами, в последнее время несколько чаще индекс Серенсена. Формула Жаккара более строгая, так как получаемые с её помощью коэффициенты соответствуют реальному сходству, тогда как формула (15) даёт сугубо относительные величины. Например, если в одном местообитании обитает какое-то количество видов, а в другом половина из них, то их сходство по формуле (14) будет равно 0,5, или 50%, т. е. будет соответствовать действительно имеющему место двукратному различию; формула (15) в этом случае даёт величину 2/s. Это не имеет решающего значения при сравнительных, анализах, так как и в том и в другом случае с помощью получаемого ряда цифр можно судить о степени сходства, однако при незначительных фаунистических различиях сравниваемых комплексов -некоторое завышение показателей, полученных по формуле Серенсена, может приводить к нивелированию различий.
Если сравнить полные ряды индексов сходства, р'ассчитанных по двум этим формулам, то легко можно видеть, что индексы, вычисленные по формуле (15), образуют более выровненную гиперболическую кривую (рис. 21, Б). Коэффициент Жаккара (рнс. 21, Л) имеет большую амплитуду числового выражения, т. е. большую дифференцирующую силу198
при средних и высоких показателях сходства (примерно от 30% и выше). Лишь при очень низких величинах сходства преимущество в этом плане имеет формула Серенсена. Формула Жаккара лежит в основе целого ряда других индексов, например группового сравнения по видам и по обилию (см. ниже).
Все прочие методы расчёта фаунистического сходства (их обзор дан в книге М. С. Гилярова, 1965) в принципе ничего не
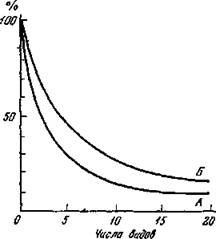
Рис. 21. Кривые, отражающие степень сходства (в °/о) при постепенном нарастании различий в видовом составе Объяснение в тексте
добавляют к тем характеристикам, которые можно получить с помощью формулы Жаккара.
Индексы фаунистического сходства весьма формальны в син- экологическом смысле. Ведь это сопоставление только видовых списков в целом, в которых разные виды могут быть крайне неравноценными в количественном отношении. Особенно осторожно их надо применять при анализе сопряженных в ландшафте группировок, так как их сходство может быть высоким в силу случайного попадания нетипичных видов. При этом степень сходства сильно зависит от полноты выборки. Эти индексы более приемлемы для фаунистических региональных анализов.
В почвенной зоологии до сих пор применялись только попарные индексы фаунистического сходства. Между тем часто возникает необходимость одновременного сравнения видового состава целой серии группировок., Ряд методов такого сравнения предложен в гидробиологии и геоботанике. Например, А. А. Са- лазкин (1971) при типологии озер по их фауне сравнивал сходство каждого сообщества с одним из них, занимающим крайнее положение по интересующему нас фактору (температуре, гумус- ности и т.
д.). Это позволяет расположить группировки в ряд по степени отношения к какому-либо фактору. Такой метод безусловно может быть использован в почвенной зоологии. Для оценки видового состава серии описаний растительности В. Д. Лопатин (1965) предлагает сразу несколько формул, из которых простейшая — отношение количества видов, общих всем сравниваемым группировкам, к числу видов, отмеченных в дан- 199ной серии описаний. Видовое сходство в пределах группы местообитаний отражает также «индекс 'биотической дисперсии» Коха (Koch, 1957):

где Т — сумма видов, отмеченных в каждом из сравниваемых списаний; 5 — число видов во всей серии описаний; п — число описаний. Примечательно, что при п = 2 эта формула превращается в формулу Жаккара.
Едва ли может быть признан удачным метод расчета индексов фаунистического сходства, при котором отбираются только виды с высокой численностью или встречаемостью (Jablonsky, 1964; Loksa, 1966). Эти коэффициенты имеют смысл лишь при использовании полных видовых списков, выявленных сравнимыми методами.
В отношении типологии группировок животного населения более показательны индексы количественного сходства, или общности по обилию, расчёт которых основан на тех же принципах, что и в формулах фаунистического сходства. Используются в основном формулы Жаккара и Серенсена, в которых значение количества видов заменяется соответствующими величинами обилия, встречаемости. В геоботанике это впервые сделал Гли- зон (Gleason, 1920), который рассчитывал сходство по встречаемости на основе формулы Серецсена. Нам представляется вполне возможным проводить подобные анализы по любым показателям обилия (численности, количество гнёзд, масса, энергетические характеристики и т. д.) на уровне любых таксонов (видов, родов, семейств, экологических групп).
Н. И. Нефедов (1930) при анализе распределения муравьев использовал отношение количества гнезд видов, общих двум сравниваемым площадкам, к суммарному количеству гнезд всех видов на двух площадках.
В. Н. Беклемишев (1961) в качестве «индекса общности по численности» предлагает отношение суммарной численности видов, общих сравниваемым единицам среды, к численности всех видов в обеих группировках.Недостатком этих формул является то, что в них не учитываются возможные существенные различия по обилию между общими видами. Более показательны методы расчета сходства по обилию с использованием не полной численности общих компонентов, а её минимальных значений (из двух по каждому виду), т. е. тех величин, которые действительно общие для сравниваемых группировок. Один из таких показателей представляет собой сумму минимальных значений удельных обилий каждого из видов, общих двум сравниваемым совокупностям (Renko- пеп, 1938 — цит. по Wallwork, 1970; Вайнштейн, 1949, 1967)
Knsl~ SSmin, (17)
200
где Лп» — коэффициент общности удельного обилия по терминологии Б. А Вайнштейна (1967); S— удельное обилие — часть или процент обилия каждого вида от суммарного обилия всех видов в группировке. Этот показатель не зависит от величины сравниваемых выборок, так как используются процентные отношения в каждой группировке. Необходимо иметь в виду специфические особенности Kns, ограничивающие его применение при сравнительных анализах животного населения. Во-первых, здесь учитывается только обилие общих видов, что может сильно нивелировать отличия сравниваемых сообществ в целом; во-вторых, не принимаются во внимание различия в общих величинах обилия двух группировок Р. Л. Наумов (1964) при расчётах сходства по обилию группировок птиц использовал формулу:

где с —меньший (из двух) показатель обилия каждого вида, а — суммарное обилие всех видов в одной группировке, Ь — то же, в другой. Как видим, она соответствует формуле фаунистического сходства Жаккара, где в числителе — количество общих видов, а в знаменателе'—сумма видов в обеих группировках минус число общих видов.
Анализ получаемых индексов показывает, что формула (18) более чувствительна и в целом дает цифры, более тонко отражающие степень количественного сходства. Так, из табл. 8 видно, что при расчете по формуле (17) ве-Таблица 8
Индексы сходства группировок дождевых червей на территории Центральночерноземного заповедника, рассчитанные по численности отдельных видов
| Группировки | А | в | в | г | Д |
| А | 100 | 34 | 30 | 63 | 24 |
| Б | 13 | 100 | 46 | 35 | 37 |
| В | 20 | 32 | 100 | 54 | 35 |
| Г | 37 | 24 | 42 | - 100 | 31 |
| д | 5 | 13 | 34 | 9 | 100 |
Условные обозначения: А — дубрава, Б — некоснмая степь, В — южный склон лога, Г — дннще лога, Д —пастбище.
Примечание. Справа от диагонали 100%-ного сходства — индексы, рассчитанные по формуле (17), слева — по формуле (18)
личины коэффициентов сильно снивелированы (максимальные и минимальные различаются менее чем в 3 раза); в левой части таблицы эти различия восьмикратные. Во всех случаях соотношение коэффициентов по формуле (18) ближе к логически пред-
201
полагаемым. Очевидно, что нрй простом суммнронаний только удельных обилий общих видов слишком велико маскирующее значение степени различия по небольшому количеству наиболее массовых компонентов.
Вместе с тем индекс К, обладающий рядом специфических особенностей, может быть использован с определенной целью, например, при анализе однотипных группировок со сходным видовым составом, равно как и при анализе на уровне крупных таксонов.А. С. Константинов (1969) рассматривает коэффициенты сходства сообществ с позиций теории множеств и приходит к выводу, что наиболее репрезентативные показатели дает расчет их по формуле Серенсена. Индекс сходства по обилию получается как отношение удвоенной суммы минимальных значений (нз каждых двух) общих видов к суммарному обилию всех видов в обоих сообществах, т. е.

Всё сказанное выше о формулах Жаккара и Серенсена относится и к этим формулам. При сравнении по обилию нескольких пар группировок, в которых обитают одни и те же виды (начиная с одного), но при 2-кратных, 4-кратных и так далее различиях в численности каждого, формула (18) даёт соответственно величины индексов 50, 25% и т. д., тогда как формула (19) в каждом случае—большие значения. При полном спектре-степеней сходства индексы по формуле (19) дают значительно более выровненную гиперболическую кривую, чем по формуле (18). В ряде случаев это обстоятельство может сильно нивелировать различия. В связи с этим нам представляется предпочтительней формула (18).
Коэффициенты сходства по обилию наиболее применимы при анализах группировок с небольшим количеством видов или при обработке данных по крупным группам. В принципе их можно вычислять по любым таксонам и в любых показателях обилия. При достаточно высоком качественном сходстве и небольшом количестве компонентов коэффициенты по обилию достаточно показательны. Однако при сравнении разнотипных группировок, включающих большое количество компонентов, применение их ограничено, так как сходство по одному — двум массовым видам, которыми часто бывают мало специфичные убиквисты, сильно маскирует степень сходства по остальным. В связи с этим Б. А. Вайнштейн (1967) предложил новый показатель — попарные коэффициенты «биоценологического сходства»[5], объ
единяющий величины сходства удельного обилия и фаунистической общности:

где Лпв — коэффициент общности удельного обилия (17), Kf — коэффициент фаунистического сходства, рассчитываемый по формуле Жаккара. Наш опыт использования этого показателя для анализа животного населения лесостепи и тундры заставляет считать его заслуживающим широкого применения в син- экологических исследованиях. Особенно хорошие результаты даёт его применение при анализе комплекса микроартропод, представленных относительно большим количеством видов. В свете сказанного выше о формулах (17) и (18), вероятно, рационально для этого использовать показатель общности по обилию, рассчитываемый по однотипной с жаккаровской формуле (18), а не 17, как это предлагает Б. А. Вайнштейн. Тогда формула принимает вид:

Как и в предыдущих случаях, нет принципиальных препятствий для расчёта коэффициента Вайнштейна как по численности, так и по массе при анализе на уровне любых таксонов.
Необходимо иметь в виду, что Кп и К/ попользуются как равноценные сомножители. В связи с этим при анализе разнотипных, но соседствующих в ландшафте группировок сходство может быть завышено в силу большого значения Kt (за счет легкости случайного обмена характерными для каждой группировки достаточно многочисленными видами). Так, в лесостепи К«, между некоторыми соседствующими, но разнотипными группировками (например, степь и лесной склон соседнего лога) в некоторых случаях оказался выше, чем между луго-степными участками и полянами. При этом Кп давал обратную картину. Очевидно, в некоторых случаях сугубо типологическое сходство лучше выявляется индексами Кп, величина которых в первую очередь определяется обилием массовых видов, что отражает наиболее характерные особенности экологических режимов.
Дальнейшее использование попарных коэффициентов может быть самым различным, например для построения экологических рядов, дихотомических схем и т. д. Для большей наглядности и удобства при последующем анализе попарные коэффициенты можно оформлять в виде диаграмм по класеам сходства (рис. 22), как это обычно практикуется в отношении индексов фаунистического сходства и сопряженности видов (Balogh, 1958; Макфедьен, 1965).
Имеется ряд способов преобразования попарных коэффици- qhtqb сходства с целью сравцитсдвцой характеристики серии
S03
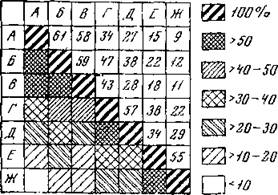
Рис. 22. Диаграмма-решетка сходства группировок почвенных беспозвоночных, учтенных в пробах 25X25 см в различных биотопах Центральночерноземного заповедника Индексы (в %) рассчитаны яа основании соотношения массы семейств беспозвоночных А — косимая степь, Б— некосимая степь; В — центральная часть поляны среди дубравы; Г — край поляны; Д — облесенный лог в степи, Е — плаксхрная дубрава; X —дннще оврага в лесу
местообитаний и их классификации по степени сходства друг с другом. Один из них предложен Маунтфордом (Mountford, 1962 — цит. по Гилярову, 1965 и Wallwork, 1970). Метод состоит в следующем. В таблице индексов попарного сходства (видов или местообитаний), например на рис. 22, находят пару, имеющую наибольшее сходство. В нашем случае (на рис. 22) это А и Б (индекс 61%). Далее рассчитываем сходство этой пары со всеми прочими местообитаниями по схеме: сходство АБ с В будет равной—58,5 (округленно 58), т. е. индексы сходства каждого члена нашей пары с В, найденные на рис. 22, складываем и делим на число слагаемых. Таким образом последовательно находим сходство АБ со всеми прочими местообитаниями, Затем с учетом вновь рассчитанных индексов и рис. 22

В этой диаграмме вновь находим максимальный индекс. Это 58 — сходство пары А Б с В. Объединяем эти три выдела и вновь по той же схеме рассчитываем сходство тройки АБВ со всеми прочими, для чего складываем сходство каждого местообитания (имея перед глазами рис. 22) с А, Б, В и делим на 3. Полученные индексы используем для составления новой диаграммы, где в одной графе будет стоять тройка АБВ. В этой диаграмме максимальный индекс будет между Г и Д. Находим сходство
204
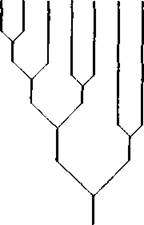
Рис. 23. Классификация группировок почвенных беспозвоночных различных биотопов Центрально-Черноземного заповедника на основании индексов количественного сходства по методу Маунтфорда (Mountford, 1962 — из Гилярова, 1965)
Длина вертикальных линий, связывающих пары, пропорциональна различию
Обозначения см. на рис. 22
Л- & в ГД ЕЖ
этой пары со всеми прочими, включая и АБВ. В последнем случае будет уже шесть слагаемых:
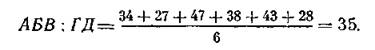
Во вновь составленной диаграмме максимальное сходство будет между Е и Ж;'в следующей — между АБВ и ГД. В результате все наши местообитания разделились на две группы: АБВГД и ЕЖ- Найдем сходство между этими группами. После этого построим диаграмму, показанную на рис. 23, в которой местообитания располагаются попарно в виде дихотомической схемы, начиная снизу с последних двух групп и далее в порядке увеличения индексов в соответствии с приведенными выше расчетами. Для большей наглядности вертикальные линии, соединяющие пары, можно откладывать в определенном масштабе, отражающем сходство (или различие — чем меньше линия, тем больше сходство). Иногда сбоку изображается принятый масштаб. В нашем простейшем примере искомая классификация четко соответствует логически предполагаемым и очевидным соотношениям.
Кроме непосредственного сопоставления попарных коэффициентов, хорошим вспомогательным показателем, позволяющим охарактеризовать ряд группировок по степени их специфичности, могут служить суммы коэффициентов по каждому из сравниваемых выделов. Наибольшую сумму имеют группировки, занимающие «среднее» положение в рассматриваемой совокупности, обнаруживающие наибольшее сходство с большинством прочих. Снижение этого показателя, который можно именовать индексом специфичности, соответствует повышению специфики условий биотопов, например, увеличению или уменьшению увлажненности. Это хорошо иллюстрируется табл. 9 (надо иметь в виду, что это данные для микрогруппировок в пределах одного зонального сообщества). Вполне естественна наибольшая специ-
205
Таблица 9
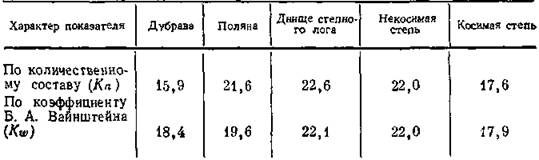
Попарные коэффициенты Б. А. Вайнштейна, в долих и их суммы для микро' .группировок коллембол пятнистой тундры Таймыра (по Ананьевой, 1971)

Условные обозначения: I — пятна голого грунта, II — пятна с фрагментарным растительным покровом, III — пятна, покрытые сплошной тонкой моховой дерниной, IV — мощная моховая дернина вокруг пятен.
фичность группировки коллембол, обитающих на голом грунте. Наоборот, некоторым «усредненным» составом характеризуются в данном сукцессионном ряду промежуточные стадии зарастания пятен. В серии группировок по всему ландшафтному профилю данного района наибольшие величины этого индекса оказались у выделов с чертами типичных зональных сообществ (Ананьева, 1971; Чернов и др., 1971). В лесостепи, наоборот, максимальные показатели тяготеют к группировкам, занимающим промежуточное положение между собственно зональными сообществами при достаточном увлажнении (поляны, некосимые участки лугостепи, в мощном травостое которых сочетаются луговые, степные и лесные элементы, а также днища логов с чрезвычайно разнообразным высокотравьем). Меньшие величины индекса получены для косимой степи и дубравы—основных зональных типов сообществ лесостепи. Небольшие различия показателей в табл. 10 отражают характерные особенности ландшафтной структуры животного населения лесостепи, где в силу относительной сглаженности градиента гидротермических режимов контраст между цлакорными зональными и интразональными группировками не столь существен, как в других зонах. При подобном анализе животного населения в тундровой зоне различия достигали трехкратной величины. Очевидно, аналогичным образом можно, использовать и коэффициенты по обилию и фауне. Прй этом нагляднее рассчитывать их в долях или процентах от общей суммы в пределах каждого анализируемого ряда (табл. 10).
В последнее время геоботаники стали применять формулы для расчета сходства одновременно целой серии выделов по количественным показателям — численности, покрытию, встречаемости (Williams, Lance, 1965; Лопатин, 1965; Василевич, 1969, и др.). gee эти методы, безусловно, представляют ицтерес и для
ада
Таблица 10
Специфичность комплексов беспозвоночных, учтенных методом ручной разборки (в процентах от суммы Величин сходства по массе в пределах каждого ряда)
геозоологических исследований. Сходство по обилию в пределах серии группировок мы предложили рассчитывать исходя из формул Жаккара и Л. Коха, использовав принцип минимальных количественных показателей (Чернов, 1971):

где /„ (следуя терминологии Л. Коха) — «индекс биотической дисперсии» по обилию; 22V— сумма количественных показателей каждого компонента во всех группировках, SJVmin — сумма минимальных обилий каждого из всех компонентов, п — число сравниваемых группировок. Например, в местообитаниях I, II, III виды а, б, в имеют обилие:
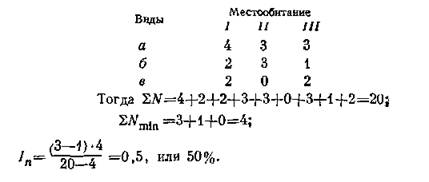
Необходимо иметь в виду, что индексы сходства могут быть крайне формальными, если в серии группировок одна-две имеют резко отличный от прочих качественный и количественный состав. Очевидно, формулу (22) имеет смысл применять лишь при достаточно большом сходстве качественного состава, в частности, при анализе по крупным таксонам. Этот индекс можно использовать для выявления различий в степени контрастности, дискретности в пределах каждого из нескольких типов сообществ (при этом могут быть использованы как средние данные
207
по отдельным топическим группировкам, входящим в состав сообщества, так и отдельные пробы). В. Д. Лопатин (1965) предлагает использовать для сравнения серии площадок двучленную формулу—произведение величин сходства по видовому составу и обилию (тот же принцип, что и в рассмотренной выше формуле Б. А. Вайнштейна).
Еще по теме СРАВНИТЕЛЬНЫЙ АНАЛИЗ ГРУППИРОВОК ПОЧВЕННЫХ БЕСПОЗВОНОЧНЫХ:
- Методы изучения пищевой избирательности беспозвоночных-сапрофагов
- Метод сравнительного анализа
- 2. Сравнительный анализ экономической эффективности
- Сравнительный анализ индивидуального и группового решения
- Сравнительный анализ особенностей философии, науки, искусства, морали
- Приложения1. Программа учебного курса «Сравнительное правоведение (основные правовые системы современности)»Общая часть Введение в теорию и историю сравнительного правоведенияТема 1. Сравнительное правоведение: метод, наука, учебная дисциплина
- 3.4 Сравнительный анализ нескольких структур
- ж) итоги сравнительного и исторического анализа
- 2.2.2 Сравнительный статистический анализ алгоритмов сглаживания
- Сравнительный анализ теоретических и экспериментальных данных
- Сравнительный анализ вольт-амперных характеристик
- § 4. Современное российское и западноевропейское правосознание в контексте формы государства: сравнительный анализ
- §3. Сравнительный анализ идеологии Хиндутвы и теории «строительства нации».
- 3.4.4. Сравнительный анализ результатов моделирования классификации экспертов
- Эмпирическая и «понимающая психология». Сравнительный анализ подходов.