Регрессивная ценопопуляция
Инвазионная ценопопуляция может перейти в нормальную, а нормальная – в регрессивную.
Возрастная структура ценопопуляции во многом определяется биологическими особенностями вида: периодичностью плодоношения, числом продуцируемых семян и вегетативных зачатков, способностью вегетативных зачатков к омоложению, скоростью перехода особей из одного возрастного состояния в другое, способностью образовывать клоны и др.
Типичный возрастной спектр называется базовым (рис. 99). Проявление всех этих биологических особенностей, в свою очередь, зависит от условий внешней среды. Меняется и ход онтогенеза, который может протекать у одного вида во многих вариантах (поливариантность онтогенеза), что сказывается на структуре возрастного спектра ценопопуляции (рис. 100).

Рис. 99. Базовый тип спектра ценонопуляций (по Л. Б. Зауголыювой, 1976) А – бурачок ленский; Б – анабазис безлистный; В – овсяница луговая; Г – типчак.
1 – базовый спектр; 2 – пределы изменения базового спектра
Разные размеры растений отражают различную жизненность особей в пределах каждой возрастной группы. Жизненность особи проявляется в мощности ее вегетативных и генеративных органов, что соответствует количеству накопленной энергии, и в устойчивости к неблагоприятным воздействиям, что определяется способностью к регенерации.
Жизненность каждой особи меняется в онтогенезе по одновершинной кривой, возрастая на восходящей ветви онтогенеза и уменьшаясь на нисходящей. У многих видов особи одного возрастного состояния в одной ценопопуляции могут иметь разную жизненность. Эта дифференциация особей по жизненности может вызываться разнокачественностью семян, разными сроками их прорастания, микроусловиями среды, воздействием животных и человека, конкурентными отношениями. Высокая жизненность может сохраняться до смерти особи во всех возрастных состояниях или снизиться в ходе онтогенеза. Растения высокого уровня жизненности нередко проходят все возрастные состояния ускоренными темпами. В ценопопуляциях часто преобладают растения среднего уровня жизненности. Одни из них проходят онтогенез полностью, а другие пропускают часть возрастных состояний, переходя перед отмиранием на более низкий уровень жизненности. Растения низшего уровня жизненности имеют сокращенный онтогенез и часто переходят в сенильное состояние, едва приступив к цветению.
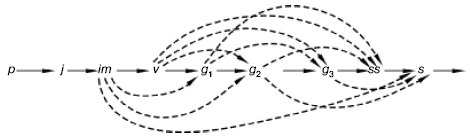
Рис. 100. Варианты развития ежи сборной в разных экологических условиях (по Л. А. Жуковой, 1985).
Латинскими буквами обозначены возрастные состояния растений, а пунктирными линиями – их возможная последовательностьОсоби одной ценопопуляции могут развиваться и переходить из одного возрастного состояния в другое с разной скоростью. По сравнению с нормальным развитием, когда возрастные состояния сменяют друг друга в обычной последовательности, может наблюдаться ускорение или задержка в развитии, выпадение отдельных возрастных состояний или целых периодов, наступление вторичного покоя, часть особей может омолаживаться или отмирать. Многие луговые, лесные, степные виды при выращивании их в питомниках или посевах, т. е.
на лучшем агротехническом фоне, сокращают свой онтогенез, например овсяница луговая и ежа сборная – с 20–25 до 4 лет, горицвет весенний – со 100 до 10–15 лет, жабрица порезниковая – с 10–18 до 2 лет. У других растений при улучшении условий онтогенез может удлиняться, как, например, у тмина обыкновенного.В засушливые годы и при усилении выпаса у степного вида овсеца Шелля отдельные возрастные состояния выпадают. Например, взрослые вегетативные особи могут сразу пополнять группу субсенильных, реже – старых генеративных. Клубне-луковичные растения безвременника великолепного в центральных частях компактных клонов, где условия менее благоприятные (хуже освещение, увлажнение, минеральное питание, проявляется токсическое действие отмерших остатков), быстрее переходят в сенильное состояние, чем периферические особи. У свербиги восточной при усиленной пастбищной нагрузке, когда повреждаются почки возобновления, молодые и зрелые генеративные особи могут иметь перерывы в цветении, тем самым как бы омолаживаясь и продлевая свой онтогенез.
У ежи сборной в разных условиях реализуется от 1–2 до 35 путей онтогенеза, а у подорожника большого от 2–4 до 100. Возможность менять путь онтогенеза обеспечивает адаптацию к меняющимся условиям среды и расширяет экологическую нишу вида.
У двух видов степного овсеца – Шелля и опушенного – в Пензенской области четко прослежено циклическое изменение возрастных спектров в многолетней динамике. В засушливые годы популяции овсецов стареют, а во влажные – молодеют. Флюктуации возрастного спектра ценопопуляций вслед за погодными условиями особенно характерны для растений поемных лугов.
Возрастной спектр может варьировать не только вследствие внешних условий, но и в зависимости от реактивности и устойчивости самих видов. К выпасу у растений разная сопротивляемость: у одних выпас вызывает омолаживание, так как растения отмирают, не достигнув старости (например, у полыни равнинной), у других – способствует старению ценопопуляции изза снижения возобновления (например, у степного вида жабрицы Ледебура).
У некоторых видов на всем протяжении ареала в широком диапазоне условий нормальные ценопопуляции сохраняют основные черты возрастной структуры (ясень обыкновенный, типчак, овсяница луговая и др.).
Такой возрастной спектр зависит преимущественно от биологических свойств вида. В нем сохраняются прежде всего соотношения во взрослой, наиболее стабильной части. Число вновь возникающих и отмирающих особей в каждой возрастной группе сбалансировано, и общий спектр оказывается постоянным до значительных изменений условий существования. Такие базовые спектры чаще всего имеют ценопопуляции видов-эдификаторов в устойчивых сообществах. Им противопоставляются ценопопуляции, относительно быстро меняющие возрастной спектр вследствие не установившихся отношений с окружающей средой.Чем крупнее особь, тем значительнее сфера и степень ее воздействия на среду и на соседние растения («фитогенное поле», по А. А. Уранову). Если в возрастном спектре ценопопуляции преобладают особи взрослые вегетативные, молодые и средневозрастные генеративные, то вся популяция в целом будет занимать более прочное положение среди других.
Таким образом, не только численность, но и возрастной спектр ценопопуляции отражает состояние и приспособленность ее к меняющимся условиям внешней среды и определяет позиции вида в биоценозе.
Еще по теме Регрессивная ценопопуляция:
- 8.5.7. Динамика ценопопуляций растений
- 7.1.3. Прогрессивность и регрессивность подоходного налога
- Регрессивное поведение
- 4. Сокращенный сложный силлогизм
- МОДИФИКАЦИИ ГЛАСНЫХ
- РЕГРЕСС
- 7.1.11. Резюме
- § 70. Ассимиляция фонем в русском языке.
- Возрастная структура популяций у растений.
- Отмирающие особи
- 8. Л. В РЛЯ - л и л’.
- 2. ФОРМЫ ПРАВЛЕНИЯ
- 1. Историческое место колониализма. В XX веке европейский колониализм вступил в последнее, пятое столетие своего существования.
- АДОРНО