ИЗМЕНЕНИЕ СОСТОЯНИЯ РЫБНОГО НАСЕЛЕНИЯ ОНЕЖСКОГО ОЗЕРА С 60- х ГОДОВ XIX СТОЛЕТИЯ ПО 2000 ГОД
Л.А. Кудерский
Учреждение Российской академии наук Институт озероведения РАН, г. Санкт-Петербург, Россия
Состояние рыбного населения Онежского озера в последние десятилетия претерпело существенные изменения.
Не сбалансированный с имеющимися запасами интенсивный промысел, избирательно использующий наиболее ценные виды рыб, в совокупности с другими формами антропогенного влияния на экосистему водоема (гидростроительство, изменение качества водной среды в результате сбросов загрязняющих веществ промышленными предприятиями, сельскохозяйственными объектами, коммунальным сектором, в недалеком прошлом молевой сплав леса и т.д., а также браконьерский лов) оказали негативное влияние на численность ряда рыб. В результате этого наблюдается антропогенная сукцессия озерного ихтиоценоза, выражающаяся в изменении соотношения между отдельными видами, снижении или даже потере промыслового значения некоторых из них. В структуре рыбного населения существенно сократилась роль озерно-речных видов и рыб с длительным жизненным циклом, которые замещаются мелкими короткоцикловыми озерными формами. В связи с этим, во-первых, сокращается товарная ценность уловов, во-вторых, возникает угроза сложившемуся биоразнообразию рыбного населения озера. Происходящие изменения в ихтиофауне неоднократно отмечались различными авторами, освещавшими как общее состояние рыбного населения, складывающееся в последний период времени, так и неблагополучное положение с отдельными видами (озерный лосось, озерная форель, палия, озерно-речные и озерные сиги, судак и т.д.). Однако в этих публикациях охватывается обычно ограниченные временные отрезки. Кроме того, они часто носят описательный характер и не содержат количественных данных, наглядно отражающих происходящие изменения.В связи с наблюдающимися существенными преобразованиями, возникает задача анализа динамики рыбного населения за возможно более длительный отрезок времени выраженной в количественных показателях.
Наиболее приемлемыми материалами для такого анализа могли бы служить данные официальной промысловой статистики, дополняемые биологическими оценками состояния популяций облавливаемых видов. К сожалению, надежные статистические сведения имеются за относительно короткий период 30-х годов XX столетия и за 1946-1990 гг. Эти данные, безусловно, должны использоваться при анализе состояния рыбного населения. Но они, во-первых, охватывают все же короткий временной промежуток и, во-вторых, характеризуют тот период, во время которого в озере осуществлялся достаточно интенсивный промысел, а его водная система уже усиленно эксплуатировалась различными отраслями народного хозяйства (лесосплав, гидроэнергетика, сброс загрязненных сточных вод и т.д.). Между тем в литературе начиная со второй половины XIX столетия содержится немалая информация, которая при соответствующей обработке могла бы использоваться для анализа состояния рыбного населения озера в целом (и отдельных видов рыб) в различные временные отрезки. В частности, имеются сводные работы по ихтиофауне озера, рыбопромысловые описания, содержащие оценки состояния уловов (и запасов) отдельных видов по водоему и по различным районам, характеристики эксплуатации отдельных видов промыслом и т.п. При использовании всего массива накопленных сведений временной отрезок анализа изменения состояния рыбного населения озера может быть увеличен примерно до 130 лет.Автор проанализировал литературные источники начиная со второй половины XIX столетия по настоящее время и имеющуюся в них информация по теме настоящего доклада сгруппировал по следующим четырем периодам: 1) 60-годы XIX столетия (Кесслер, 1868; Данилевский, 1875; Пушкарев, 1900); 2) 30-е годы XX столетия (Покровский, Смирнов, 1932; Покровский, 1939, 1947); 3) вторая половина 50-х - первая половина 60-х годов XX столетия (Александров и др., 1959; Гуляева, Кудерский, 1964; Лузанская, 1965; Рыбы Онежского озера..., 1983; Биологические основы..., 1984); 4) 90-е годы XX столетия (Красная книга Карелии, 1995; Кудерский и др., 1997; Китаев, 1999; Современное состояние., 2000).
Отличительная черта многих использованных публикаций - качественная оценка (и часто подробная) состояния описанных рыб. Для преобразования имеющегося массива качественных и количественных характеристик в единую систему показателей использовался балльный метод, изложенный в предыдущих работах (Кудерский, 1989; 1996; Кудерский, Печников, 2002) и являющийся разновидностью экспертных оценок. В соответствии с этой методикой состояние каждого вида рыб в озере характеризовалось индексом, состоящим из двух цифр (баллов) соединенных знаком плюс, каждая из которых может принимать значения от 1 до 5. Левая цифра отражает распространенность вида в озере и качественную оценку его обилия, правая - промысловое значение и мероприятия по регулированию промысла и искусственному воспроизводству запасов. При расположении первых цифр по горизонтали и вторых по вертикали получается поле значений индексов, охватывающих весь диапазон возможной изменчивости состояния популяций рыб в данном водоеме. При этом индекс, характеризующий состояние вида, находится на пересечении указанных строк и столбцов. Виды рыб, находящиеся в удовлетворительном состоянии, характеризуются минимальными значениями индекса, находящиеся в критическом или близком к нему состоянии - максимальными. В итоге все поле возможных значений индекса состояния видов может быть представлено следующей матрицей (табл. 1). Общая степень благополучия состояния видов понижается при переходе от левого верхнего угла матрицы к правому нижнему. Она наиболее удовлетворительная при значениях индекса близких к 1+1 и наименее при значениях, приближающихся к 5+5. Образуя подобные индексы для каждого вида по выделенным выше четырем периодам можно в количественной форме выразить изменения во времени состояния каждого конкретного вида, а также всего рыбного населения озера в целом.
Таблица 1
Индексы состояния видов рыб
| Численность и распространенность вида | Промысловое значение вида, меры по регулированию промысла и охране запасов | ||||
| 1. - не промысловый, охранные меры не требуются. | 2. - промысловый, охранные меры не требуются. | 3. - промысловый, достаточны «Правила рыболовства». | 4. - промысловый, необходимо искусственное воспроизводств о запасов. | 5. - состояние критическое, что отражается занесением в «Красную книгу». | |
| 1. Широко распространенный, многочисленный. | 1 +1 | 1 +2 | 1 +3 | 1 +4 | |
| 2. Широко распространенный, малочисленный. | 2+1 | 2+2 | 2+3 | 2+4 | 2+5 |
| 3. Ограниченно распространенный, многочисленный. | 3+1 | 3+2 | 3+3 | 3+4 | |
| 4. Ограниченно распространенный, малочисленный. | 4+1 | 4+2 | 4+3 | 4+4 | 4+5 |
| 5. Редкий, исчезающий | 5+1 | 5+2 | 5+3 | 5+4 | 5+5 |
Таблица 2
Индексы состояния видов и форм рыб Онежского озера по периодам
| Вид рыб | 60-е годы XIX века | 30-е годы XX века | Вторая половина 50-х - первая половина 60-х годов XX века | 90-е годы XX века |
| Минога речная | 2+2 | 2+2 | 2+2 | 2+2 |
| Лосось озерный | 1 +2 | 1 +4 | 1 +4 | 4+5 |
| Форель озерная | 2+2 | 2+2 | 1 +4 | 4+5 |
| Палия | 3+2 | 3+2 | 3+3 | 4+5 |
| Ряпушка | 1 +2 | 1 +2 | 1 +2 | 1 +3 |
| Сиги озерно-речные | 1 +2 | 1 +4 | 2+4 | 4+5 |
| Сиги озерные | 1 +2 | 1 +3 | 1 +3 | 1 +3 |
| Хариус | 2+2 | 2+3 | 2+3 | 2+3 |
| Корюшка | 1 +2 | 1 +2 | 1 +2 | 1 +2 |
| Щука | 1 +2 | 1 +2 | 1 +3 | 1 +3 |
| Плотва | 1 +2 | 1 +2 | 1 +2 | 1 +2 |
| Елец | 2+2 | 2+2 | 2+2 | 2+2 |
| Язь | 1 +2 | 1 +2 | 1 +2 | 1 +3 |
| Гольян | 2+1 | 2+1 | 2+1 | 2+1 |
| Пескарь | 4+1 | 4+1 | 4+1 | 4+1 |
| Уклея | 2+2 | 2+2 | 2+2 | 2+2 |
| Густера | 2+2 | 2+2 | 2+2 | 2+2 |
| Лещ | 1 +2 | 1 +2 | 1 +2 | 1 +3 |
| Карась | 4+2 | 4+2 | 4+2 | 4+2 |
| Голец усатый | 2+1 | 2+1 | 2+1 | 2+1 |
| Сом | 5+2 | 5+2 | 5+2 | 5+5 |
| Угорь | 2+2 | 4+2 | 5+5 | 5+5 |
| Налим | 1 +2 | 1 +2 | 1 +2 | 1 +2 |
| Колюшка 3-х иглая | 1 +1 | 1 +1 | 1 +1 | 1 +2 |
| Колюшка 9-ти иглая | 1 +1 | 1 +1 | 1 +1 | 1 +2 |
| Судак | 1 +2 | 1 +2 | 1 +3 | 1 +3 |
| Окунь | 1 +2 | 1 +2 | 1 +2 | 1 +2 |
| Ерш | 1 +2 | 1 +2 | 1 +2 | 1 +2 |
| Подкаменщик пестроногий | 4+1 | 4+1 | 4+1 | 4+5 |
| Подкаменщик | 2+1 | 2+1 | 2+1 | 2+5 |
Первый этап выполняемого анализа вспомогательный.
Он заключается в составлении уточненного списка видов рыб, зарегистрированных в каждом из выделенных четырех периодов. Общее число видов и форм, вовлеченных в анализ за почти 130-летний отрезок времени принимается равным 30. Для всех них установлены значения индекса состояния в соответствии с изложенным выше методическим приемом. Значение полученных индексов для каждого вида и формы по четырем периодам приведены в табл. 2.Второй этап анализа сводится к характеристике изменения индекса состояния по каждому виду. Он дает исходный материал для последующей оценки состояния рыбного населения озера в целом. Как видно из табл. 2, из 23 теоретически возможных вариантов индекса состояния видов и форм, отраженных в табл. 1, фактически для Онежского озера установлено 16. Распределение числа видов и форм по периодам по каждому значению индекса состояния представлено в табл. 3. Очевидно, что в 60-е годы XIX столетия все 30 видов и форм не требовали выполнения каких либо охранных или рыбоводных мероприятий. Спустя 70 лет в 30-е годы XX столетия не нуждались в таких мероприятиях 26 видов и форм, еще через примерно 30 лет в конце 50-х - начале 60-х годов XX столетия таких видов и форм осталось 21. Наконец, еще примерно через 40 лет в 90-е годы XX столетия число подобных видов и форм сократилось до 15.
Таблица 3
Число видов и форм рыб Онежского озера с конкретным значением индекса состояния (по периодам)
| Баллы | 1 | 2 | 3 | 4 | 5 | Итого |
| 60-е годы XIX столетия | ||||||
| 1 | 2 | 13 | - | - | - | 15 |
| 2 | 3 | 7 | - | - | - | 10 |
| 3 | - | 1 | - | - | - | 1 |
| 4 | 2 | 1 | - | - | - | 3 |
| 5 | - | 1 | - | - | - | 1 |
| Итого | 7 | 23 | - | - | - | 30 |
| 30-е годы XX столетия | ||||||
| 1 | 2 | 10 | 1 | 2 | - | 15 |
| 2 | 3 | 5 | 1 | - | - | 9 |
| 3 | - | 1 | - | - | - | 1 |
| 4 | 2 | 2 | - | - | - | 4 |
| 5 | - | 1 | - | - | - | 1 |
| Итого | 7 | 19 | 2 | 2 | - | 30 |
| Вторая половина 50-х - первая половина 60-х годов XX столетия | ||||||
| 1 | 2 | 8 | 3 | 2 | - | 15 |
| 2 | 3 | 4 | 1 | 1 | - | 9 |
| 3 | - | - | 1 | - | - | 1 |
| 4 | 2 | 1 | - | - | - | 3 |
| 5 | - | 1 | - | - | 1 | 2 |
| Итого | 7 | 14 | 5 | 3 | 1 | 30 |
| 90-е годы XX столетия | ||||||
| 1 | - | 7 | 6 | - | - | 13 |
| 2 | 2 | 4 | 1 | - | 1 | 8 |
| 3 | - | - | - | - | - | - |
| 4 | 1 | 1 | - | - | 5 | 7 |
| 5 | - | - | - | - | 2 | 2 |
| Итого | 3 | 12 | 7 | - | 8 | 30 |
Таким образом, в настоящее время, спустя 130 лет после начала отсчета изменений состояния рыбного населения Онежского озера, в нем в специальных мерах поддержания численности не нуждается только 50% видов и форм, против исходных 100%. При этом темпы роста числа видов и форм, требующих выполнения различных мер по охране и искусственному воспроизводству, возросли.
Если за первые 70 лет только четыре вида стали нуждаться в специальных мерах охраны и воспроизводства, то спустя еще 60 лет это число возросло еще на 11 видов и форм. Отмеченное явление оказывается следствием нарастающего антропогенного «давления» на экосистему озера.Приведенные выше индексы состояния видов (табл. 2) допускают выполнение над ними ряда операций, с помощью которых возможно получение количественных показателей, обобщенно отражающих изменения в рыбном населении озера на протяжении анализируемых отрезков времени.
Этот прием - третий этап анализа изменения рыбного населения озера. Одной из операций служит получение индекса состояния осредненного (абстрактного) вида, рассчитанного для рыбного населения озера в целом. Вычисление индекса состояния осредненного вида осуществляется следующим образом: для определенного периода времени суммируются все левые (соответственно - правые) цифры индексов состояния по каждому виду и форме и полученная сумма делится на показатель общего числа видов и форм (в нашем случае - 30), составляющих по исходному условию рыбное население озера. В итоге получается искомый индекс состояния некоего абстрактного вида - среднего применительно ко всему рыбному населению. Эти индексы состояния осредненного по всему рыбному населению озера вида оказываются равными:
- 60-е годы XIX столетия................................................................... 1,8+1,8
- 30-е годы XX столетия.................................................................... 1,9+2,0
- вторая половина 50-х - первая половина 60-х годов XX столетия....... 1,9+2,3
- 90-е годы XX столетия.................................................................... 2,2+2,9
Уже сами величины индексов состояния осредненного (абстрактного) вида отражают происходящие в рыбном населении озера неблагоприятные изменения. Так, увеличение левой цифры индекса, отражающей распределение и численность вида, с 1,8 в первом периоде до 2,2 в четвертом (то есть в 1,2 раза) свидетельствует о некотором ухудшении этого показателя: о сокращении распределения и уменьшении численности осредненного вида для озера в целом. Точно также увеличение правой цифры индекса состояния осредненного вида с 1,8 в первом периоде до 2,9 в четвертом (то есть в 1,6 раза) свидетельствует о существенном снижении промысловой значимости осредненного вида и возрастании усилий, необходимых для выполнения мероприятий по охране и искусственному воспроизводству численности.
Еще белее наглядно происходящие изменения в рыбном населении отражаются следующим показателем состояния осредненного вида, выражаемым в условных единицах и вычисляемым по формуле:
C = (N+L+R)/ 3
где C - показатель состояния осредненного вида для рыбного населения озера в целом, N - % видов для которых индекс состояния не изменился при переходе от одного промежутка времени к другому, L - средняя левая цифра индекса состояния вида в % к такой же средней для последующего периода времени, R - средняя правая цифра индекса состояния вида в % к такое же средней последующего периода времени. Сумма N, L, R делится на три. Этот показатель удобен в связи с тем, что выражает единой цифрой изменения трех ведущих характеристик состояния осредненного вида при переходе от одного периода времени к другому: индекса состояния, распределения и обилия, промысловой значимости и усилий, необходимых для охраны и воспроизводства численности.
Из структуры этой формулы вытекают следующие общие оценочные выводы: 1) максимальное значение C не превышает 100; 2) по мере увеличения величин левой и правой цифр индекса состояния вида (по сравнению с исходным состоянием) значения L и R уменьшаются, а с ними уменьшается и величина C; 3) уменьшение величины показателя состояния осредненного вида свидетельствует о негативных тенденциях в динамике рыбного населения, причем чем меньше величина C, тем сильнее изменено рыбное население по сравнению с исходным состоянием. Соответствующий расчет по приведенной выше формуле дает следующие величины для показателя состояния осред- ненного вида для рыбного населения озера в целом (в условных единицах): первый период - 100,0; второй - 89,3; третий - 81,0; четвертый - 62,4. Негативные изменения между состоянием рыбного населения Онежского озера в 60-е годы XIX столетия и в 90-е годы XX выступают достаточно выпукло и количественно отражают сложившееся неблагополучное положение в рыбной составляющей экосистемы озера, что неоднократно отмечалось в литературе.
Для более определенных выводов о глубине произошедших изменений в рыбном населении озера целесообразно установить возможные (теоретические) пределы колебания значений показателя состояния осредненного вида и положение в этом диапазоне фактического значения C. С этой целью примем следующие граничные данные: 1) состояние всех 30 видов и форм рыб подверглось изменениям; в таком случае величина N=0; 2) левая и правая цифры индекса состояния каждого вида и формы достигли предельного значения 5; в этом случае L=36,0 и R=36,0. Отсюда по приведенной выше формуле C=24,0. Таким образом, теоретически возможные изменения величины C для рыбного населения Онежского озера лежат в пределах от 100,0 при неизменном состоянии рыбного населения в начале и конце рассматриваемого периода до 24,0 при максимально возможных изменениях. В этом случае современное фактическое значение C=62,4 находится на расстоянии 37,6 условных единиц от неизменного (исходного) состояния и 38,4 условных единицы от минимально возможного. Если принять, что изменения C на всем диапазоне значений 100,0-24,0 носят линейный характер, то допустимо считать, что в настоящее время степень изменения рыбного населения озера находится где то на половине пути между стационарным (исходным) состоянием и полной катастрофой, понимая под последней максимальное изменение индексов состояния всех обитающих в озере видов и форм рыб.
В заключение отметим, что примененный в настоящей работе переход от качественных характеристик, содержащихся в традиционных ихтиологических описаниях, к количественным с помощью балльной методики может быть использован для аналогичных целей и для других водоемов при наличии разнесенных по времени данных по состоянию рыбного населения.
СУТОЧНОЕ ВЕРТИКАЛЬНОЕ РАСПРЕДЕЛЕНИЕ БИОМАССЫ КОПЕПОД СЕТНОГО ЗООПЛАНКТОНА В КАНДАЛАКШСКОМ ЗАЛИВЕ БЕЛОГО МОРЯ В РАЗНЫЕ СЕЗОНЫ И.П. Кутчева, Д.М. Мартынова
Учреждение Российской академии наук Зоологический Институт РАН, г. Санкт-Петербург, Россия e-mail: kartesh@gmail.com
Введение
Картина распределения биомассы зоопланктона по сезонам в различных районах Белого моря на разных глубинах представлена в работах Эпштейн (1957, 1963), Перцовой (1970, 1971), Прыгун- ковой (1977). Большинство работ по изучению вертикального распределения зоопланктона в Белом море проводилось в светлое время суток. Однако миграции зоопланктона могут накладывать большой отпечаток на распределение биомассы (Виноградов, 1968). Данные о том, как она меняется на разных глубинах в течение суток, и как миграции зоопланктона влияют на этот показатель в разные сезоны, практически отсутствуют. Цель данной работы - изучить суточную динамику распределения биомассы копепод сетного зоопланктона в Белом море в летний и осенний периоды.
Материалы и методы
Исследования проводились 1-2 июля и 28-29 сентября 1998 г. на двух суточных станциях в районе островов Илейки в Кандалакшском заливе Белого моря (66°22.7' N, 33°49.2' E). Глубина здесь составляет 100 м (максимальная глубина Белого моря 343 м). Материал собирался двумя сетями Джеди с различным диаметром ячеи фильтрующего конуса (0.1 и 0.2 мм) по стандартным горизонтам 0-10 м, 10-25 м, 25-50 м, 50-100 м. Сборы были сделаны через каждые 3--4 часа и приурочены к приливно-отливному циклу (8 парных серий, 128 проб). Количественная обработка зоопланктонных проб проведена по стандартной методике (Руководство..., 1980).
В данной работе рассмотрена динамика суточного вертикального распределения биомассы основных видов ракообразных сетного зоопланктона в период исследований. Изучали динамические характеристики девяти видов: Calanus glacialis, Metridia longa, Pseudocalanus minutus, Oncaea
borealis, Oithona similis, Microsetella norvegica, Acartia longiremis, Temora longicornis, Centropages ha-
matus. Учитывались все возрастные стадии, включая науплиальные.
Результаты и обсуждение
В летний период по биомассе доминировал P. minutus во всех горизонтах до 50 м в сборах обеими сетями в течение суток (60-80% всей биомассы). В слое 50-100 м главенствовала M. longa в дневное время (30-40%), а ночью и здесь P. minutus вышел на первое место (52-56%), т.к. часть M. longa поднялась вверх и ее биомасса в нижнем горизонте уменьшилась в два раза.
Осенью картина не столь однозначна. Прежде всего, наблюдались различия по составу зоопланктона и его биомассе в слое 0-10 м как в разное время суток, так и в сборах разными сетями. Днем в пробе крупноячеистой сетью доминировала по биомассе T. longicornis (57%), а в пробе, взятой более частой сетью - O. similis (39%). Ночью их вытеснил P. minutus в сборах обеими сетями (48 и 54% соответственно). В слое 10-25 м днем доминировал P. minutus (42 и 32% соответственно), хотя в пробе мелкоячеистой сетью биомассы O. similis и P. minutus практически равны (по 32 %). Ночью доминировала M. longa, особенно в сборах крупноячеистой сетью, до 88% всей биомассы. В слое 25-50 м днем преобладал P. minutus (73 и 56% соответственно), ночью - M. longa (80 и 70%). В слое 50-100 м в течение суток доминировал P. minutus (68 и 66% днем, 86 и 78% ночью).
Науплиусы отмечены во всей толще воды. Роль науплиальных стадий копепод более значительна в летний период. Относительно общей биомассы копепод их значение максимально в слое 10-25 м (до 10%), а не в слое 0-10 м (до 3%). Глубже они составляли до 2% биомассы и менее. В сентябре науплиусов больше в слое 0-10 м (до 3%), в слое 10-25 м в среднем за сутки 1,4%, глубже их роль в биомассе несущественна.
Картина суточной динамики распределения общей биомассы копепод по горизонтам была схожа в летний и осенний периоды (рис. 1, 2). Заметное увеличение биомассы копепод произошло как в июле, так и в сентябре в вечернее и ночное время во всех горизонтах до 50 м. В июле в слое 0-10 м биомасса копепод увеличилась к 22 ч. в полтора раза, что совпало с фазой полной воды. К 2 ч. она уже снизилась (фаза середины отлива), но превышала дневные показатели и почти не изменилась к 5 ч. утра (фаза малой воды). Далее вновь последовало увеличение значений биомассы. В слое 10-25 м биомасса копепод увеличивалась более продолжительный период времени. Ее значения были высокими в период с 22 до 4 ч. ночи и превосходили дневные значения более чем в два раза к 2 ч. ночи. В слое 25-50 м биомасса копепод увеличилась в период с 19 до 24 ч., максимально в 22 ч. - более чем в полтора раза. А вот в слое 50-100 м биомасса копепод уменьшилась в период с 19 до 4 ч. Особенно это заметно по сборам сетью с ячеей 0.2 мм в 2 ч., почти в два раза. Таким образом, во всех горизонтах отмечено наибольшее изменение величины биомассы в 22 ч., и только в слое 10-25 м она еще более увеличилась и в 2 ч. ночи.
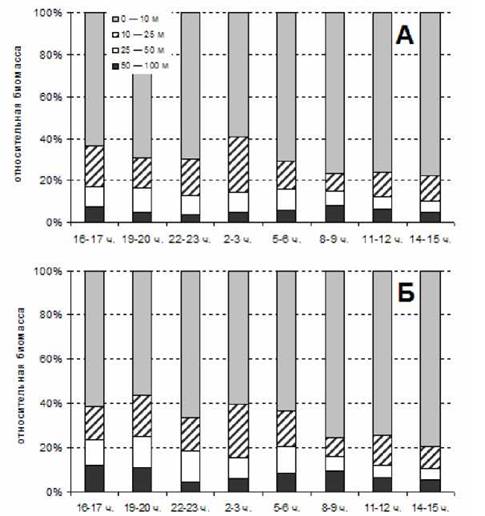
Рис. 1. Динамика биомассы копепод на суточной станции 1-2 июля 1998 г. А - диаметр ячеи 0.1 мм; Б - диаметр ячеи 0.2 мм
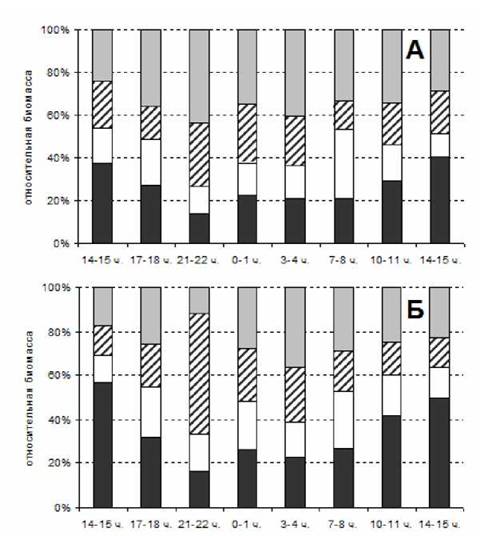
Рис. 2. Динамика биомассы копепод на суточной станции 28-29 сентября 1998 г. А - диаметр ячеи 0.1 мм; Б - диаметр ячеи 0.2 мм. Обозначения как на рис. 1
В осенний период в слое 0-10 м заметное увеличение биомассы началось с наступлением сумерек в 19 ч. и держалось более полусуток. Максимально - более чем в 3.3 раза в сборах сетью с ячеей 0.2 мм и в 2.6 раза - сетью с ячеей 0.1 мм в 3 ч. ночи, что совпало с фазой полной воды, как и летом. В слое 10-25 м увеличение биомассы копепод наблюдалось меньший промежуток времени, с 21 до 5 ч. Однако здесь в 22 ч. наблюдалось почти десятикратное увеличение величины биомассы при сборах сетью с ячеей 0.2 мм и почти трехкратное - сетью с ячеей 0.1 мм. В слое 25-50 м величина биомассы также начала увеличиваться с наступление сумерек, и превышала дневные значения более 12 часов, причем максимальное превышение (в 4 и 3 раза соответственно) наблюдалось в 7 ч. утра, очевидно, когда животные уже опускались. И в слое 50-100 м отмечено уменьшение значений биомассы с 18 ч. до 7 ч.утра.
Изменения биомассы произошли благодаря перемещениям копепод. Летом в ночное время в поверхностных слоях увеличилась численность P. minutus, это совпало с фазой полной воды. А осенью в слой 10-25 м в вечернее и ночное время поднялась в массе M. longa из нижних горизонтов, за счет этого резко увеличилась биомасса. При этом общая численность зоопланктона в сентябре мало изменилась в отличие от июльских сборов. В июле ночью биомасса M. longa стала больше в слое 25-50 м, чем в слое 50-100 м. Небольшое количество самок (1%) попалось в слое 10-25 м (что не отразилось на величине биомассы) и не отмечено выше в отличие от осенних миграций. Осенью днем метридия держалась в слое 50-100 м, ночью 10% всей популяции поднялось в слой 0-10 м и 10% осталось в слое 50-100 м, большая часть особей поднялась во второй горизонт.
Выводы
Таким образом, наибольшие изменения биомассы копепод отмечены в слое 10-25 м как в летний, так и в осенний сезоны. Особенно это проявилось осенью за счет подъема M. longa, что несомненно связано с периодом осеннего размножения этого вида. Во всех горизонтах до 50 м произошло увеличение биомассы копепод в ночное время, и только в слое 50-100 м она уменьшилась. В слое 0-10 м заметна связь с фазами прилива.
Возможными механизмами, вызывающими суточные вертикальные миграции копепод, могут быть, с одной стороны, преимущественная фитофагия большинства из них (Перуева, 1984; Martynova et al., unpublished) и, таким образом, стремление к верхним, наиболее богатым фитопланктоном слоям воды. С другой стороны, стремление избежать как поимки их зрительными планктонофагами (Виноградов, 1968), так и окислительного стресса (Martynova, Gordeeva, in press) заставляет рачков мигрировать в светлое время суток в глубокие слои воды.
Литература
Виноградов М. Е., 1968. Вертикальное распределение океанического зоопланктона М.: Наука. М. 319 с.
Перуева Е. Г., 1984. Сравнительная оценка питания двух массовых копепод Белого моря // Океанология. Т. 24. С. 813-818.
Перцова Н. М., 1970. Зоопланктон Кандалакшского залива Белого моря // Тр. ББС МГУ. Т. 3. С. 34-45.
Перцова Н. М., 1971. О количественном вертикальном распределении зоопланктона в Кандалакшском заливе Белого моря // Комплексные исследования природы океана. Изд. МГУ. Вып. 2. С. 152-162.
Прыгункова Р. В., 1977. Зоопланктон Кандалакшского залива в сравнении с зоопланктоном на декадной станции Д-1 в губе Чупа (Белое море) // Экология морского планктона. Исслед. фауны морей. Х1Х(ХХУП). Л.: Зоологический институт. С.100-108.
Руководство методам биологического анализа морской воды и донных отложений, 1980. / Ред. А.В. Цыбань Л.: Гидрометеоиздат. 191 с.
Эпштейн Л. М., 1957. Зоопланктон Онежского залива и его значение в питании сельди и молоди рыб // Материалы по комплексному изучению Белого моря. Т. 1. Изд. АН СССР. М.-Л. С. 315-349.
Эпштейн Л. М., 1963. Зоопланктон Белого моря и его значение в питании сельди // Проблемы использовния промысловых ресурсов Белого моря и внутренних водоемов Карелии. Вып. 1. Изд. АН СССР, М.-Л. С. 98-104.
Martynova D. M., Gordeeva A. V. Light-dependent behaviour of major zooplankton species in the White Sea with some implications to their feeding and life cycle patterns, Journal of Plankton Research, in revision.
Еще по теме ИЗМЕНЕНИЕ СОСТОЯНИЯ РЫБНОГО НАСЕЛЕНИЯ ОНЕЖСКОГО ОЗЕРА С 60- х ГОДОВ XIX СТОЛЕТИЯ ПО 2000 ГОД:
- 1.3. «Естественное движение» населения на Ставрополье в конце 20-х - начале 30-х годов XX века и отражение в переписях 1937 и 1939 годов изменений в демографической сфере ставропольского села, произошедших в годы коллективизации.
- IV. Состояние науки уголовного права к началу шестидесятых годов XIX в.
- Ранние империи (2000 год до н.э. — 600 год н.э.)
- Глава З Психологические портреты шестилетних и семилетних детей 90-х годов XX столетия
- Исследования Восточной Африки в конце 70-х — начале 80-х годов. Путешествие Томсона к Великим озерам и открытие им озера Руква
- 2000 год. OZON.ru с новым владельцем
- Склонность XIX столетия к изучению истории
- Заключение. Литературные традиции XVIII столетия и русская литература XIX века
- 16. Изменения в общей части уголовного законодательства конца 20-х - начала 30-х годов. Изменения в законодательстве о государственных и имущественных преступлениях.
- Государственный строй Италии в последней трети XIX - начале XX столетия.
- 1.2. Миграции населения Ставропольского и Терского округов на рубеже 20-х - 30-х годов ХХ века.
- 1966 год – изменение стратегии лунной гонки?