КРУГОВОРОТ ВЕЩЕСТВ II СОЛЕВОЙ РЕЖИМ РАСТИТЕЛЬНОСТИ ПУСТЫНЬ [108]
Для различных растительных сообществ среднеазиатских пустынь, были определены: 1) общий запас сухой органической фитомассы, включая надземную и подземную (в большинстве случаев до глубины 50 см}· части; 2) количество ежегодного опада за счет отмирающих надземных частей и корней; 3) общее количество ежегодно поступающих в почву азота и зольных элементов.
Расчет величины 2 для однолетних растений трудности не представляет, в данном случае она равна величине 1. У многолетних же видов нельзя непосредственно определить массу ежегодно отмирающих корней. Эта доля исчислена исходя из средней продолжительности жизни многолетних видов. Например, для Artemisia badghysi, достигающей в среднем· 20-летнего возраста, ежегодный опад принимался равным V20 части всего количества многолетних корней. Для травянистых многолетних злаков было принято, что ежегодно отмирает V3 общей массы корней. Эти расчеты· оказались сходными с данными непосредственного определения корневых
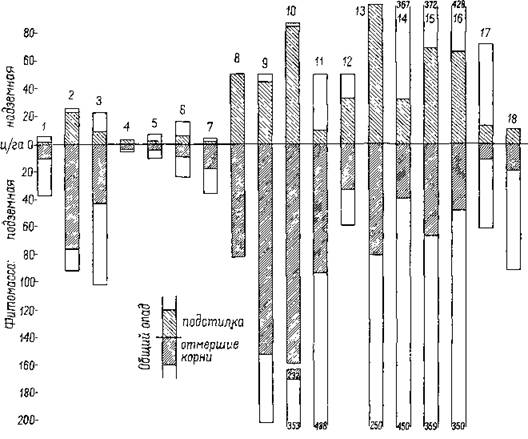
Рис. 164. Фитомасса (общий запас сухой органической массы) и ежегодный опад (надземный и подземный) в различных растительных сообществах среднеазиатской пустыни. Указаны общее количество опада (оп.) в % фитомассы, содержание азота (N) и зольных элементов в % опада £(по Родину).
1 — ассоциация Artemisiakemrudi—Anabasis salsa (on. 27%, N 1,5%, зольность 5,1 %);
2 — Artemisia badghysi с эфемерами (on. 86,%, N 0,9%, зольность 4,7%); 3 — чистая заросль Anabasis salsa (on. 42%, N 2,3%, зольность 9,3%); 4 — однолетние галофиты с Suaeda (он. 85%, N 1,3%, зольность 13,4%); 5 — чистая заросль Halocnemum Strobilaceum (оп. 30%, N 1,8%, зольность 16,5%); в — Suaeda совместно с однолетними галофитами (оп. 40%, N 1,6%, зольность 11,2%); 7—чистая заросль Anabasis aphylIa (оп.
49%, N 2,8%, зольность 6,5%); S — чистый луг с Eremopyrum Orientale на намытой лаве в предгорье (оп. 100%, N 1,2%, зольность 7,6%, много SiO2); 9 —луг с Arfemisia и Poabulbosa в западине (оп. 77%, N 1,3%, зольность 6,9%); 10 — ассоциация Aeluropus—Vicia—Alopecurus в низине на луговом сероземе (оп. 73%, N 1,7%, зольность 5,2%, много SiO2); 11 —заросли Haloxylon aphyllum с участием Tamarix (1 %) над солеными грунтовыми водами (оп. 19%, N 1,9%, зольность 6,3%); 12 — кустарниковые тугаи (Tamarix—Aeluropus) над солеными грунтовыми водами (оп. 61%, N 1,1%, зольность 13%); 13 — тростниковое болото (Phragmi- tes) в дельте Амударьи; надземная фитомасса может достигать максимально IOQ т/га (оп. 51%, N 1,5%, зольность 11 %’, 58% золы составляет SiO2, 10% Gl, 7,5% S, 14% К, 6% Ga, 2,5% Na); 14 — пойменный лес с Salix Songarica—Calamagrostis pseudophrag- mites (on. 9%, N 0,6%, зольность 5,4%); 15 — пойменный лес с Elaeagnus Orientalis— Alhagi persarum (оп. 18%, N 1,4%, зольность 4,8%); 16 — пойменный лес с Populus diversifolia—Halimodendron halodendron, поверхность почвы слабо засолена (оп. 14%, N 1,4%, зольность 5,6%); 17—пойменный лес с Populus diversifolia—Aeluropus, засоление более заметно (оп. 17%, N 1,0%, зольность 6%); 18 — Alhagi persarum на заброшенном орошаемом участке; когда запас влаги в почве израсходован, его замещает Anabasis aphylla', корни исследованы лишь до глубины 25 cat (оп. 30%, N 1,9%,зольность 5%).
масс в степных сообществах. Для Poa bulbosa, ежегодно образующего множество отмирающих эфемерных корешков, для расчета было принято, что •ежегодно в опад поступает половина общей массы корней. Всего Родиным изучено 31 растительное сообщество. Это единственные в своем роде данные, имеющиеся для области пустынь. Поэтому мы остановимся на них подробнее. На рис. 164 графически изображены общий запас сухой органической массы, надземная и подземная фитомасса в различных сообществах пустынной растительности, а также количество ежегодно уходящего в онад общего запаса органического вещества, слагаемого из массы отмерших надземных частей и корней.
Приведено также процентное содержание азота и золы в опаде. В общем, из имеющихся данных можно сделать следующие выводы:1. У всех растительных сообществ пустыни доля подземной фитомассы значительно превосходит надземную. Разница невелика только у сообществ эфемеров или у сообществ, обитающих на переувлажненных почвах. В среднем на корни приходится 75% (67—91 %) от общего количества органического вещества, в пойменных лесах — 50% (45—54%).
2. В структуре ежегодного опада наибольшую массу дают также корни. Среди пустынных формаций саксаульники занимают первое место по степени концентрации количества живого вещества в подземных органах растений.
3. Отмершая масса корней бедна золой. Основная доля зольных элементов содержится в опаде зеленых ассимилирующих органов листьев, ■стеблей. На последние приходится 9—28% всего опада; лишь при хорошей обеспеченности водой в пойменных лесах эта доля возрастает до 41—56%.
4. Чрезвычайно высокой зольностью отличаются злаки и галофиты. У первых это обусловлено содержанием SiO2 (окремневший эпидермис), у вторых связано с накоплением солей в клеточном соке растений.
5. Накопление главнейших элементов по-разному осуществляется у трех групп галофитов: а) аккумулируют преимущественно NaGl, ■б) накапливают относительно много сульфатов и в) содержат Na больше, чем это соответствует эквивалентному количеству СГ и SO^. Следовательно, натрий присутствует в клеточном соке в виде солей органических кислот.
6. Особую группу образуют галофиты, выделяющие соли. В образуемый ими опад поступает относительно небольшое количество элементов, но известно, что виды Tamarix и Aeluropus сбрасывают некоторое неучитываемое количество соли на поверхность почвы в течение всего вегетационного периода.
7. Содержание азота в опаде пустынных сообществ относительно высокое. Доля азота в опаде в среднем составляет: в пустыне — 1,7% (в пойменных лесах — 0,9%), в степях — 1,2 и в бореальных лесах — 0,5%.
8. Содержание золы в опаде пустынных формаций (8%) превосходит аналогичные показатели для опада лесных (3,3%) и степных (7,3%) сообществ.
9. Чем больше общий запас органической массы, тем меньшая ее доля поступает в опад. В лесах органическое вещество как бы консервируется в одревесневших стволе и корнях и снова вовлекается в круговорот веществ лишь спустя длительное время. Напротив, в сообществах эфемеров, создающих сравнительно небольшую фитомассу, опад ежегодно полностью минерализуется. Еще интенсивнее совершается биологический круговорот в сообществах такырных водорослей. Косвенным путем определена их сухая масса в расчете на 1 га площади; она составила 0,5—0,6 т. Можно предполагать, что в течение вегетационного периода кругооборот веществ в этих своеобразных биоценозах водорослей совершается неоднократно.
Доля высших растений на такырах чрезвычайно мала. На слабозарос- щих такырах общая фитомасса высших растений достигает 0,15—0,3 тіга, на сильнозаросших — увеличивается до 0,6—1,6 т!га. Опад в этом случае составляет 100%, так как живыми остаются только семена; зольность опада равна 7,3—9,1%, содержание азота в нем достигает 2%. В заключение приведем составленную Родиным по различным источникам таблицу^ в которой сопоставлены между собой различные группы формаций по размерам фитомассы на единицу площади и по количеству ежегодного опада (табл. 46).
Таблица 46
Биологический запас (общее количество органической массы) и количество ежегодного опада в различных группах
формаций (в т/г)
| Группы формаций | Биологический запас | Опад |
| Сосновые леса .................................... | 200 | 7 |
| Еловые леса......................................... | 150—380 | 6-20 |
| Липовые смешанные леса .... | 160-220 | 5—10 |
| Дубовые леса ...................................... | 160-500 | 6-27 |
| Осинники ............................................ | 190-250 | 12-25 |
| Остепненные луга............................... | 11-16 | 6-9 |
| Луговые степи..................................... | 20—23 | 9-13 |
| Разнотравно-типчаково-ковыль- | ||
| ные степи...................................... | 18—34 | 8-14 |
| Пустынные сообщества Централь- | ||
| ной Азии: | ||
| Древесные тугаи................................. | 70-80 | 7—13,5 |
| Кустарниковые тугаи ........................ | 6-11 | 3,5-6,5 |
| Травяные тугаи................................... | 35 | 18 |
| Пустынные луга.................................. | 24-45 | 20—32 |
| Саксауловые пустыни......................... | 55 | 10,5 |
| Солянковые пустыни......................... | 2-12,5 | 1-5,5 |
| Эфемерово-полынные пустыни | 11—14 | 6-10,5 |
| Полынные пустыни ........................... | 4,5 | 1,2 |
| Зарастающие такыры ......................... | 0,6-1,6 | 0,6-1,6 |
| Водорослевые такыры с немно- | ||
| гими эфемерами.......................... | 0,1-0,3 | 0,1-0,3 |
Для пустынь характерно повсеместное распространение засоленных почв. Заметную роль в образовании засоленных почв играет отложенио пыли. В больших количествах соль содержится во всех морских осадочных отложениях, и прежде всего в часто обнажающихся на поверхности морских третичных породах. Переотложение солей происходит обычно с помощью воды; соли смываются на ниже расположенные участки рельефа и интенсивно накапливаются в местах испарения капиллярной влаги. Растения, произрастающие на засоленнйх местообитаниях, по лощаюг вместе с водой соли, совершенно не нужные им для обмена веществ, такие, как хлористый натрий и сернокислый натрий. Эти соли накапливаются в транспирирующих органах растений, которые, как известно, одновременно осуществляют процесс фотосинтеза. Поэтоа у такие органы характеризуются наивысшей зольностью, и прежде всего содержат много балластных солей (NaCl, Na2SO4). Для большинства растений такая аккумуляция солей даже в небольших размерах оказывается вредной; поэтому на засоленных почвах эти виды не выдерживают конкуренции. Галофиты, напротив, легко переносят относительно большую концентрацию солей; соли в известной мере даже стимулируют их рост. Эти виды успешно развиваются на засоленных почвах и со временем начинают дажо
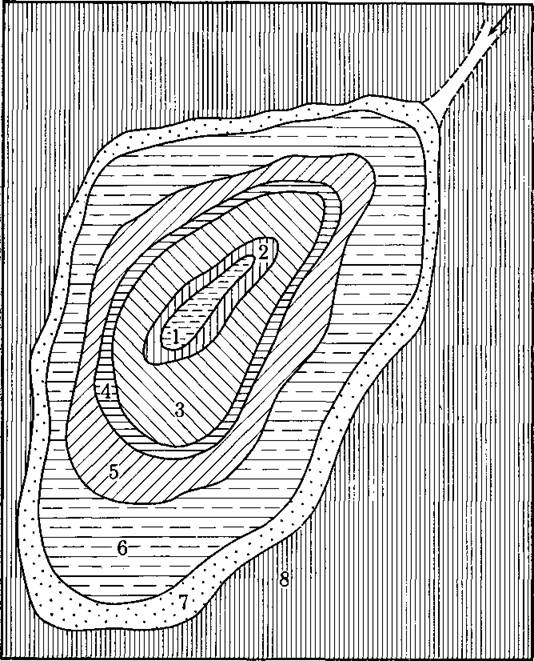
Рис. 165. Схема распределения растительности в засоленной озерной депрессии в долине реки Чиж, западный Казахстан (по Ларину).
1 — вода; 2 — заросли тростника; 3 — Agropyrum repens или Calamagrostis (Gala- tella glabra)', 4 —галофиты (Halocnemum, Suaeda, Atriplex, реже Aeluropus); 5 — Alro- pis C Agropyrum Cristatum или Galatella Villosa', в — то же с Artemisia incana (г;ло- фильное растение); 7 — Atriplex Verrucifera', 8 — комплексная полупустыня. Западина обладает, по-видимому, некоторым подземным стоком, иначе засоление увеличивалось бы по направлению к центру, как это отражено на рис. 166.
доминировать, поскольку быстрее растущие, но менее солеустойчивые виды по мере нарастания соленосности почвы один за другим выбывают мз’конкурентной борьбы. Вокруг засоленных впадин наблюдается очень резко выраженная поясность растительности, обусловленная активизацией соленакопления в направлении к центру впадины (рис. 165 и 166). Там, где «одержание солей становится предельно высоким, растительность полностью отсутствует. На этот факт мы неоднократно указывали в т. I и отмечали различия в количестве и качестве солей, поглощаемых растениями, которые пронизывают своими корнями одну и ту же почву. Мы различаем
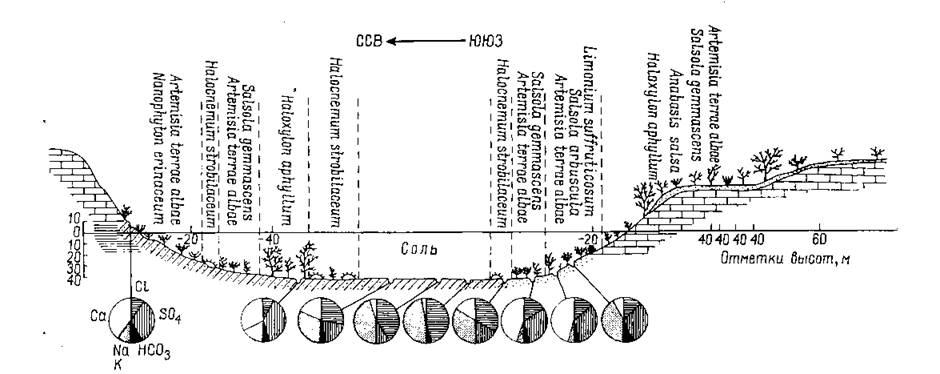
Рис. 166. Схема распределения растительности по поперечному профилю засоленной впадины Каунды (по Викторову и др.).
Вертикальный масштаб увеличен в 20 раз. В кружках показан химический состав (в %) водных вытяжек из почвы (места взятия проб отмечены на профиле ямками). В центре впадины наблюдается присутствие только NaGl, к периферии увеличивается доля сульфатов и в некоторой степени также растворимых карбонатов.
виды, накапливающие в своих транспирирующих органах преимущественно NaCl и, следовательно, являющиеся суккулентами, и виды, которые аккумулируют наряду с хлоридами сравнительно много сульфатов и поэтому отличаются меньшей суккулентностью или вовсе суккулентами не являются (см. т. I, стр. 386, 405, 433). Эти рассуждения подтверждаются данными Родина, осуществившего многочисленные определения состава золы для растений среднеазиатских пустынь.
Из его весьма содержательных материалов мы приведем здесь только часть анализов и ограничимся сравнением транспирирующих и ассимилирующих органов. Эту роль исполняют либо^листья, либо, при редукции последних, фотосинтезирующие однолетние побеги; те и другие ежегодно возобновляются (рис. 167, 168).
На графиках представлены только сумма зольных веществ (зольность) в процентах от сухого веса и содержание тех элементов, которые в виде легкорастворимых солей накапливаются в клеточном соке и обусловливают повышение осмотического давления. Из анионов — это ионы хлора и сульфата, из катионов — натрий; калий приведен лишь в тех случаях, когда он составляет более 5% общего количества золы. Содержание Ca и Mg на графиках не отражено. Правда, эти элементы также могут существовать в растворе, но большая их часть, видимо, присутствует в нерастворимой форме; так, например, кальций, на долю которого часто приходится более 10% и даже более 20% золы, преимущественно содержится в растениях как щавелевокислый кальций, то есть в форме конечного продукта обмена веществ.
У злаков общее количество зольных элементов вследствие окремнения эпидермиса обычно резко повышено, что может создать впечатление о высоком содержании солей в листьях злаков. Поэтому на графиках всякий раз это окремнение отмечено. Для негалофитов показано также содержание фосфора, имеющее в определении кормовой ценности растений особое значение. При выпасе скота на естественных пастбищах следует учитывать, что фосфор является элементом, который находится в растениях в минимальном количестве. У галофитов содержание фосфора по сравнению с другими зольными компонентами всегда очень мало.
В работе Родина зольность выражена как процентное содержание Na2O, K2O, Cl, SO3. Для удобства сравнения содержаний катионов и анионов, возможно, было бы целесообразнее выразить эти значения в грамм-эквивалентах. Однако пробный расчет показал, что общая картина графика изменилась бы мало. Столбцы для сульфата оказались бы несколько короче, чем для хлора. В меньшей мере это относится и к К по сравнению с Na. Относительная длина столбцов для Cl в сравнении с длиной столбцов Na осталась бы почти такой же. Поэтому от пересчета мы отказались. Бросается в глаза, что столбцы анионов у галофитов часто бывают длиннее столбцов катионов. Во всех случаях, когда содержание калия для достижения равновесия недостаточно, следует предполагать, что в клеточном соке имеются ионы кальция. -"I 7? 7^
Для многих галофитов длина столбца натрия намного больше длины столбцов хлора и сульфата. В этих случаях в клеточном соке должны присутствовать значительные количества органических кислот. При озолении образуется карбонат натрия, и зола будет отличаться резко щелочной реакцией. То же самое произойдет при разложении опада. Следовательно, опад таких видов, минерализуясь, может вызвать щелочное содовое засоление и солонцеватость почв. Поэтому мы называем эти виды щелочными галофитами, хотя в живых органах клеточный сок, несомненно, имеет кислую реакцию. На рис. 167 приведены данные анализов золы галофитов. Мы подразделяем их на галофиты, накапливающие соли, и галофиты, выделяющие соли. Последние через особые железки

Рис. 167. Состав золы галофитов по данным Родина.
а — вычислено на золу без SiO2', содержание SiO2 в золе 65%; б,— то же, но содержание SiO2 в золе 36%. Объяснения см. в тексте.

Рис. 168. Состав золы негал офитов по данным Родина. Объяснения см. в тексте.
в листьях могут сбрасывать некоторое количество поглощенных солей (прежде всего NaCl). Поэтому содержание в них золы оказывается существенно ниже. У Tamarix выделившаяся соль еще долго остается на поверхности листьев. Неясно, учитывалось ли это обстоятельство при определении средней зольности. Возможно, что относительно высокое содержание NaCl у Tamarix Iaxa, так же как и в последней пробе Aeluropus, объясняется именно этой особенностью солевого режима.
В отношении галофитов, не выделяющих соль, обращает на себя внимание крайне высокая зольность отдельных видов, иногда достигающая 1Z3 всей массы сухого вещества; при этом соли калия занимают долю, намного меньшую, чем соединения натрия. Лишь в Anabasis aphylla и Salsola rigida они составляют около половины. Иначе обстоит дело с анионами. На их основе можно выделить три типа галофитов, связанных между собой переходными типами:
р-1) хлоридные галофиты, накапливающие много хлоридов и мало сульфатов;
16-0434
2) сульфатные галофиты, аккумулирующие относительно много сульфатов;
3) щелочные галофиты, в золе которых хлориды и сульфаты присутствуют в сравнении с содержанием натрия в небольшом количестве.
Поскольку растения в первую очередь освобождаются от NaGl, в золе выделяющих соль видов Tamarix и Aeluropus наблюдается относительно высокое содержание K2O и SO3.
Совершенно иную картину дает анализ золы негалофитов (рис. 168), которые лишь в крайнем случае растут на слабозасоленных местообитаниях. Содержание золы только у солеустойчивых Matricaria и Phragmites превышает 10%. Однако у последнего зола на 69% состоит из SiO2- Без SiO2 зольность Phragmites окажется ниже 10% — величина, близкая к содержанию золы у Stipa.
Некоторое количество NaCl поглощают и эти растения, обычно больше Cl, чем Na. Содержание последнего в золе существенно меньше концентрации калия; в большинстве случаев хлора в ней также меньше, чем SO3. Однако следует учитывать, что часть SO3 связана с серой, которая содержится в белковых соединениях. Листья типичных видов древесных тугаев, связанных с пресными грунтовыми водами, ведут себя как листья настоящих негалофитов.
В заключение еще раз подчеркнем, что все изложенное выше относится лишь к транспирирующим органам растений. Содержание растворимых неорганических солей в осевых и корневых органах даже у типичных галофитов весьма и весьма мало.
Еще по теме КРУГОВОРОТ ВЕЩЕСТВ II СОЛЕВОЙ РЕЖИМ РАСТИТЕЛЬНОСТИ ПУСТЫНЬ [108]:
- Солевой режим.
- Биогеохимические круговороты вещества в биосфере
- ПРОДУКТИВНОСТЬ И КРУГОВОРОТ ВЕЩЕСТВ В АРКТИЧЕСКОЙ ЗОНЕ
- Глобальный биогеохимический круговорот вещества на планете
- Особенности водно-солевого обмена.
- 3.4. Особенности выживания в пустыне
- 1. Стадия Человека Примитивного является переходной между состоянием живого вещества и разумного вещества
- Выживание в жаркой пустыне.
- КВАДРАТИК ПЕРВЫЙ. Стакан воды в пустыне
- T аримская впадина и пустыни T акла-Макан (Кашгария)
- 3.1.2. Условия превращения растительных остатков
- 26. ТРЕБОВАНИЯ В ОБЛАСТИ ОХРАНЫ ОКРУЖАЮЩЕЙ СРЕДЫ ПРИ ИСПОЛЬЗОВАНИИ РАДИОАКТИВНЫХ ВЕЩЕСТВ, ЯДЕРНЫХ МАТЕРИАЛОВ, А ТАКЖЕ ХИМИЧЕСКИХ ВЕЩЕСТВ В СЕЛЬСКОМ И ЛЕСНОМ ХОЗЯЙСТВЕ
- Биогеохимический круговорот воды.