ГЕОГРАФИЯ БИОӯ РАЗНООБРАЗИЯГЕНОГЕОГРАФИЯ
Популяционная генетика, зароӯдившаяся как ветвь прикладной математики в начале XX стоӯлетия, исследует динамику наследственной информации в поӯколениях. Мерой такой информации служит гетерозигот ность, т.
е. разнообразие генов, унаследованных от родителей потомством и определяющих различия особей внутри попуӯляции. Например, на надкрыльях божьих коровок легко обӯнаружить строго индивидуальные, наследуемые пигментные узоры (пятна, точки), а популяционную принадлежность рыӯбок пецилий, излюбленного объекта аквариумистов, можно определять по рисунку на хвостах у самцов. Серин работ были выполнены после обнаружения методами цитологии индиӯвидуальных различий и в строении хромосом (например, по наличию или отсутствию инверсий). Такого рода генетически детерминируемая, индивидуальная прерывистая изменчиӯвость носит название полиморфизма и определяется как наӯличие в одном и том же местообитании двух или более дискӯретно отличающихся внутривидовых форм в таких количе ственных соотношениях, что самая редкая из них не может поддерживаться лишь давлением повторяющихся мутаций.10В
ГЕОГРАФИЯ БИОРАЗНООБРАЗИЯ
Однако многие виды внешне единообразны, мономорф- ны, — их гены скрыты от наблюдателя и, следовательно, генеӯтический анализ невозможен. Как преодолеть такого рода затӯруднение? Развитие молекулярной биологии позволяет теперь изучать генетику любого вида, опираясь на скрытые наследӯственные различия, «записанные» в белковых структурах оргаӯнизма. Эти особенности выявляются специальными процедураӯми: либо иммунологическими тестами, либо при электрофореӯзе белков. Точно такой же полиморфизм у самых различных видов обнаружен за последние 10-15 лет при изучении первичӯной структуры ДНК, как ядерной, так и неядерной (например, митохондриальной), так что арсенал технических средств поӯпуляционной генетики необыкновенно расширился. Вместе с тем продолжительность и высокая стоимость ДНК-тестов пока еще ограничивают их широкое применение.
Напротив, белковый полиморфизм легко выявляется при сравнительно умеренных денежных затратах на оборудование и реактивы, а общепринятая методика позволяет работать с массовым маӯтериалом (сотни образцов за день) и используется во многих лаӯбораториях. Суть методики в следующем: кровь или экстракӯты различных тканей от отдельных растений или животных помещают в специальные аппараты и в нейтральных мелкопоӯристых поддерживающих средах (гелях) сортируют молекулы в электрическом поле по заряду или другим признакам, детерӯминируемым наследственностью. Оказалось, что особи внутри популяции различаются составом крови, а также некоторыми особенностями строения определенных белков, и такие отличия передаются неизменными от родителей потомкам.Достаточно посмотреть на рис. 8, где между особями тиӯхоокеанского лосося-нерки видны различия, определяемые двухаллельным геном, кодирующим синтез фермента фосфо- глюкомутазы.
Таким образом, с точки зрения генетики, популяция — смесь генотипов. Многие биологические особенности популяӯций, прежде всего численность и продуктивность, зависят от этой их генетической структуры. Если соотношения генотиӯпов, а точнее, частоты генов, в популяции неизменны в поӯколениях, то популяция устойчива. Если соотношения меняӯются, то говорят об изменяющейся популяции. За время, проӯшедшее с конца 1960-х гг., когда метод электрофореза белков стал широко использоваться в популяционных исследовани-
109
ГЛАВА Ш
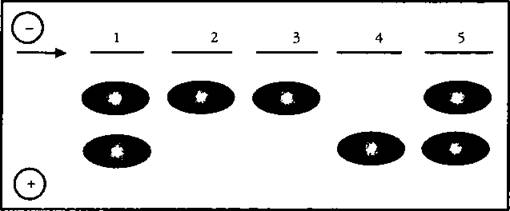
Рис. 8.
Наследственный полиморфизм фермента фосфоглюкомутазы из мышечной ткани тихоокеанского лосося-нерки, Oncorhynchusnerka: 1,5 — гетерозиготы АВ; 2-4 — гомозиготы АА и ВВ, соответственно; электрофорез в крахмальном геле; стрелка указывает стартовую позицию; направление миграции молекул в электрическом поле — от катода к аноду. Расшифровка схемы такова: один ген — одна полоса на электрофореграмме; разными номерами обозначены разные рыбы из одной и той же популяции..
У рыб 2, 3 выявляется один ген, у 4 — другой, у 1 и 5 представлены оба гена, это — гетерозиготы, т. е. типы, проӯмежуточные между двумя первыми, гомозиготными генотипамиях, биохимическая наследственная изменчивость открыта уже у более чем 2000 различных видов — от микроорганизмов до человека.
Изучение десятков генов, разных по локализации и функӯции, позволило вскрыть гигантский объем биохимического полиморфизма видов и уровней гетерозиготности — показаӯтеля, жизненно важного для организма. Оказалось, что около трети генов, кодирующих синтез белков, имеют разные (алӯлельные) наследуемые состояния, т. е. полиморфны. А уроӯвень индивидуальной гетерозиготности равен 10-12%. Если перенести эти оценки на геном в целом (равно 50 тыс. струкӯтурных генов), то окажется, что около 15 тыс. функционируюӯщих генных локусов полиморфны, а «средний» индивидуум геӯтерозиготен по нескольким сотням своих генов. Гетерозиготность организма — не что иное, как его информационная емкость, поддающаяся строгой количественной оценке шенноновской мерой количества информации. Степень полиморфизма разӯлична у разных групп организма (табл.4).
Обычно на практике полиморфной считают популяцию с частотой гетерозигот по некоторому локусу более 1—5%.
110
ГЕОГРАФИЯ БИОРАЗНООБРАЗИЯ
Таблица 4
Генетическая изменчивость в природных популяциях неко торых основных групп животных и растений (по Левонтину, 1993)
| Организмы | Число видов | Среднее число мест обитания видов | Средний уровень полиморӯ физма | Средняя гетерози- готность |
| Беспозвоночные Дрозофила | 28 | 24 | 0,529 | 0,150 |
| Осы | 6 | 15 | 0,243 | 0,062 |
| Другие насекомые | 4 | 18 | 0,531 | 0,151 |
| Морские беспозвоночные | 14 | 23 | 0,439 | 0,124 |
| Наземные улитки | 5 | 18 | 0,437 | 0,150 |
| Позвоночные Рыбы | 14 | 21 | 0,306 | 0,078 |
| Амфибии | 11 | 22 | 0,336 | 0,082 |
| Рептилии | 9 | 21 | 0,231 | 0,047 |
| Птицы | 4 | 19 | 0,145 | 0,042 |
| Млекопитающие | 30 | 28 | 0,206 | 0,051 |
| Растения Самоопыляющиеся | 12 | 15 | 0,231 | 0,033 |
| Перекрестмо- опыляющиеся | 5 | 17 | 0,344 | 0,078 |
| Общие средние Беспозвоночные | 57 | 22 | 0,469 | 0,134 |
| Позвоночные | 65 | 24 | 0,247 | 0,060 |
| Растения | 17 | 16 | 0,264 | 0,046 |
Анализ полиморфных признаков — ключ к изучению генеӯтических процессов в популяциях, в связи с чем ранние исӯследования такого рода были выполнены на видах с хорошо выраженным полиморфизмом внешнего облика особей. Наӯкоплено множество данных о феноменологии, генетическом контроле и механизмах поддержания полиморфизма у разных видов.
По мнению Ю.П. Алтухова (1989), такого рода изменӯчивость представляет собой относительно редкое явление, имеет очевидное приспособительное значение, во многих слуӯчаях поддерживается в сбалансированной форме за счет адапӯтивного преимущества гетерозигот со столь значительными коӯэффициентами отбора, что они практически не оставляют ме-111
ГЛАВА III
ста эффектам случайного дрейфа генов, в ряде случаев ее слеӯдует рассматривать (из-за ограниченной фазы стабильности) как свидетельство прошедшей и продолжающейся на наших глазах дивергенции популяций в статус новых видов, она отӯражает запас экологической пластичности за счет постоянноӯго выщепления и комбинации различных генотипов.
Главной единицей внутривидовой дифференцировки является популяция. Согласно С.И. Коржинскому (1892), каждая раса, или, как сказали бы сейчас, географическая поӯпуляция, характеризуется определенными морфологическиӯми признаками и особым ареалом. Рассмотрим в качестве приӯмера различные популяции песца. Нормальная окраска белоӯго песца типично покровительственная: зимой снежно-белая, а летом буровато-серая, с коричневыми тонами на спине и желтоватыми на боках. В популяциях песца, обитающих в тундре на материке Евразии и на островах, соединенных зиӯмой с материком, преобладают белые особи, однако в небольӯшом количестве (0,01—1,0%) встречаются и голубые. Голуӯбой песец и зимой, и летом окрашен примерно одинаково в дымчато-серые или буроватые тона. На островах, не имеюӯщих связи с материком, голубая окраска становится основной, а белая переходит в разряд отклонений. На Шпицбергене, в Исӯландии, на небольшом о. Кильдине, отделенном от материка незамерзающим проливом шириной 2—3 км, количество гоӯлубых песцов составляет 30—80%.
Сезонная белая окраска облегчает белому песцу и защиту, и, главным образом, нападение на мелких грызунов и птиц, которые являются его основной пищей в материковой тундре. Поэтому выход больших количеств голубого песца на простоӯры материкового ареала пресекается отбором.
На островах отӯбор в пользу голубых форм происходит потому, что здесь беӯлая окраска в значительной степени теряет свое приспособиӯтельное значение. На изолированных островах резко ослаблена межвидовая конкуренция, так как здесь песец — единственӯный наземный хищник. Однако главное состоит в изменении способа добычи пищи. Островной песец питается в основном продуктами моря (выброшенная морем рыба и млекопитаюӯщие, беспозвоночные на отливе и т.д.). В этих условиях на первый план выступают положительные особенности голубоӯго песца, и он начинает господствовать над белыми формами, которые переходят в разряд отклонений.112
ГЕОГРАФИЯ БИОРАЗНООБРАЗИЯ
Другой пример. Как известно, в популяциях большинства видов моллюсков встречаются особи с прямо противоӯположными по закрученности раковинками. Если у типичных особей данного вида раковинка закручена направо, по ходу часовой стрелки, то у уклоняющихся форм она закручена налеӯво, против часовой стрелки. Частота уклоняющихся раковиӯнок может быть различной у разных видов и в разных популяӯциях одного и того же вида. Так, распространение право- и левозавитковых форм у моллюсков, обитающих в окрестносӯтях Алма-Аты, где преимущественно встречаются правозавит- ковые особи. Ограничение распространения левозавитковых форм, по-видимому, обусловлено тем, что они по сравнению с правозавитковыми обладают большей интенсивностью обӯмена веществ и меньшей способностью к ассимиляции пищи. Эти особенности лимитируют распространение левоӯзавитковых форм в Семиречье, так как здесь из-за частых и длительных засух регулярно наступают периоды голоӯдания моллюсков.
На этих примерах видно, что в процессах экологической дивергенции рас на первый план .выступает адаптация к лоӯкальным условиям среды. Как указывал В.Н. Сукачев (1928), популяции сосны представлены рядом экологических рас соӯответственно условиям обитания. Чем южнее происхождение сосны, тем хуже она переносит низкие температуры. Так, температуры, безвредные для скандинавского экотипа, губиӯтельны для среднеевропейских сосен. Оказалось, что основӯная причина этих различий — биохимические особенности тканей деревьев.
У Drosophila pseudoobscura, распространенной в западных районах Северной Америки, 27 электрофоретически различӯных вариантов ксантиндегидрогеназы были обнаружены в 146 независимых линиях и 26 различных вариантов фермента эсӯтераза были найдены в 106 линиях. У людей лишь система HLA достигает такого молекулярного разнообразия. Следоваӯтельно, люди имеют несколько сниженную молекулярную изӯменчивость, но не настолько, чтобы их выделять среди друӯгих существ. В этом сношении они типичные млекопитающие.
113
Еще по теме ГЕОГРАФИЯ БИОӯ РАЗНООБРАЗИЯГЕНОГЕОГРАФИЯ:
- 2.4. Философские проблемы географии 2.4.1. Место географии в генетической классификации наук и ее внутренняя структура
- 61. География населения как ветвь социально-экономической географии
- РЕСУРСЫ ИНТЕРНЕТ ПО СОХРАНЕНИЮ БИОӯРАЗНООБРАЗИЯ
- 3.4. Закон Био — Савара
- § 4.5. ЗАКОН БИО-САВАРА-ЛАПЛАСА
- Получение био- и энтеросорбентов из природных алюмосиликатов
- I УРОВНИ БИОӯ РАЗНООБРАЗИЯСИСТЕМНАЯ КОНЦЕПЦИЯ БИОРАЗНООБРАЗИЯ
- ТЕХНОГЕННЫЕ КАТАСТРОФЫ - УГРОЗА БИОӯРАЗНООБРАЗИЮ
- VIII МОНИТОРИНГ БИОӯ РАЗНООБРАЗИЯМЕЖДУНАРОДНЫЕ ПРОГРАММЫ МОНИТОРИНГА БИОРАЗНООБРАЗИЯ
- 05.95 по делу № 5/36, согласно которого иск АКБ «Аяр» к АООТ «Био-машприбор» о взыскании задолженности по 11 кредитным договорам
- 6. Политическая география