МОРФОБИОЛОГИЧЕСКИЕ АДАПТАЦИИ ГОРБУШИ В БЕЛОМ МОРЕ
Е.А. Дорофеева
Учреждение Российской академии наук Зоологический институт РАН, г. Санкт-Петербург, Россия
e-mail: salmo@zin.ru
История акклиматизации лососевых в географически удаленных водоемах знает много примеров неудачных интродукций, которые не привели к натурализации этих рыб.
В связи с этим, большой интерес для науки представляют случаи удачных вселений, сопровождающихся возникновением стойких самовоспроизводящихся популяций, достигающих промысловой численности.Попытки акклиматизации горбуши в разных регионах мира предпринимались неоднократно. Однако результаты их противоречивы. Так, на американском континенте было проведено вселение горбуши в бассейны рек Атлантического океана (Lear, 1975). Полная натурализация произошла только после интродукции нечетной линии горбуши в Великие Озера в 1955 г., где она образовала живую форму. Вселение четной линии положительных результатов не дало (Hoard, 1991).
В России о возможности акклиматизации России на Европейском севере высказывался еще в 19 в. Карл Бэр, что было обусловлено бедностью ихтиоценозов в этом регионе, а также постепенным падением численности самого ценного вида - семги.
Горбуша была избрана в качестве акклиматизанта в силу своих уникальных среди лососевых особенностей. Этот вид имеет самый короткий двухлетний жизненный цикл и самый высокий темп роста и массонакопления (за один год нагула в море горбуша достигает массы 1.2-3 кг). Нерест проходит в значительно более ранние сроки, чем у семги. После выхода из бугров молодь задерживается в пресной воде на короткое время (1-2 мес.) Последние две особенности в значительной мере выводили горбушу из конкуренции с семгой. У горбуши снижен «инстинкт дома» (хоминг), что позволяет ей заселять в пределах ареала многочисленные реки, в том числе те, в которых популяции семги по какой-либо причине утрачены. У горбуши есть уникальная среди лососевых особенность - образовывать репродуктивно изолированные поколения смежных лет, т.е.
заходящих на нерест в четные и нечетные годы. Каждая из этих линий имеет ряд характеризующих ее экологических и генетических отличий и, таким образом, формально эти линии ведут себя как самостоятельные виды. При этом четное поколение характеризуется в естественном ареале более поздним заходом на нерест и более низкой численностью.Кариологические данные и генетическая структура популяции горбуши свидетельствуют о большей специализации четной линии по сравнению с нечетной (Гордеева, 2003; Dorofeyeva et al., 2004). Кроме того, согласно генетическим исследованиям, высказано мнение о значительных различиях между локальными стадами горбуши (Алтухов, 1974), которые подтверждается и наблюдениями морфобиологических различий между ними (Воловик, 1967; Иванков, 1967; Енютина, 1972).
Акклиматизация горбуши в бассейне Белого и Баренцева морей насчитывает более 50 лет. Массовые перевозки икры горбуши с Дальнего Востока из южной части естественного ареала, преимущественно из рек Сахалина, начались в 1956 году и продолжались с перерывами до 1980 года. В эти же годы проводилась инкубация икры от уже образовавшихся местных производителей. В первые годы после вселения горбуша начала осваивать огромный ареал. Производители стали заходить в реки от побережья Норвегии до Печоры. К сожаления, несмотря на высокие возвраты горбуши в первые годы после интродукции (в 1960 году 76,3 тыс. экз., в 1973 - 143,6 тыс.экз., в 1975 - 103,9 тыс.экз., в 1977 - 107,7 тыс.экз.), в последующие годы началось падение ее численности и лишь в некоторые, в основном нечетные годы, подход производителей превышал 2-3 тыс.экз., а в четные годы практически был равен нулю.
Было установлено, что в основе неудач интродукции горбуши с Сахалина на Европейский Север России было значительное замедление темпа оогенеза молоди в новом ареале (Сакун, 1965; Пахомова, Хлевная, 1978; Персов и др., 1983). У молоди беломорской горбуши, в отличие от сахалинской, в период миграции в море большинство ооцитов не достигали состояния превителлогенеза, а находились преимущественно на этапе ранней профазы мейоза.
Замедление раннего гаметогенеза приводило к более позднему началу периода вителлогенеза ооцитов у самок и определяло их возврат на нерест в такие сроки, при которых температурные условия были неблагоприятными для развития молоди. Таким образом, именно температурные условия в новом ареале явились основным фактором, препятствующим акклиматизации горбуши сахалинской популяции. С 1985 года начался второй этап акклиматизации. Была завезена икра горбуши нечетной линии из Магаданской области, т.е. из северной более суровой по температурным условиях части естественного ареала. В результате численность горбуши в нечетные годы значительно увеличилась (уловы варьировали от 34 до 156 тыс. экз.). Завезенная в 1998 году из той же Магаданской области икра горбуши четной линии к увеличению численности производителей не привела (Рис. 1). В связи с этим в настоящее время есть основание рассматривать нечетную линию магаданской популяции горбуши в качестве успешного донора для интродукции в бассейн Белого моря, который нашел в новом ареале условия для нагула и воспроизводства, что с очевидностью подтверждает представления о наличии генетически обособленных локальных стад горбуши в бассейне Тихого океана. Следует отметить также, что судить об эффективности акклиматизации горбуши в Белом море возможно лишь при рассмотрении данных второго этапа интродукции (т.е. с 1985 г.), поскольку предыдущее вселение из южных районов ареала было явно неудачным, и совместное рассмотрение их может привести к ошибочным выводам. Это же относится к эффективности вселения четной и нечетной линии горбуши, поскольку только последняя дала положительный результат.На основании изучения особенностей беломорской горбуши (в основном использовались материалы о нечетной линии) с применением разных методов исследования, проведенных в 20012007 гг. на выбранных в качестве модельных реках Кереть и Умба, были получены данные, свидетельствующие об адаптивных сдвигах по ряду морфобиологических признаков: линейные размеры, масса тела, плодовитость, особенности нерестовой миграции, состояние пластических, меристиче- ских и остеологических признаков, состояние половых желез у производителей и молоди (Dorofeyeva et al., 2004; Дорофеева и др.
2005, 2007; Алексеев и др., 2005, 2008). В данной работе подведены итоги проведенного исследования.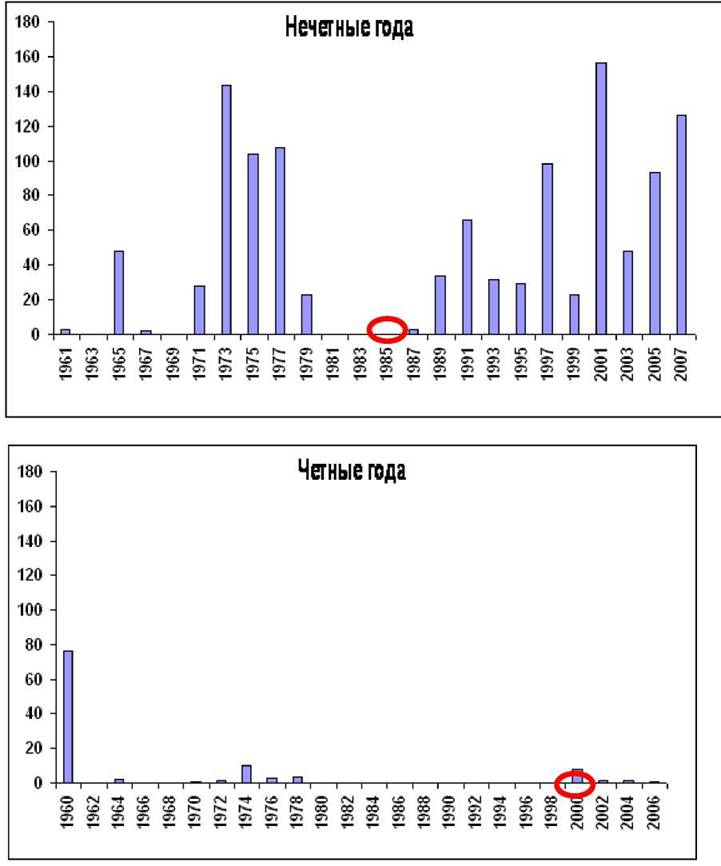
Рис.. 1. Уловы горбуши в бассейне Белого моря (тыс. экз.)
При сравнении некоторых морфобиологических признаков горбуши из бассейна Белого моря с показателями горбуши из нативного ареала (река Ола , Магаданская обл.) отмечены существенные различия (Табл. 1).
Таблица 1
Морфобиологические признаки горбуши рек Ола (Магаданская обл.) и Кереть (респ. Карелия, Белое море)
| Признак | р. Ола * | р. Кереть |
| длина взрослых самок, см 2001 г. 2003 г. | 47,3 ± 0,5 | 46,9 ± 0,35 46,8 ± 0,33 |
| вес взрослых самок, кг 2001 г. 2003 г. | 1,10 ± 0,06 | 1,41± 0,04 1,34 ± 0,32 |
| сроки ската молоди | начало мая - конец июня | начало июня - середина июля |
| сроки массовой нерестовой миграции взрослых рыб (нечетное поколение) | 15 июля - 5 августа | 10 июля - 10 августа |
| количество дней с нулевой температурой воды | 150 | 180 |
| пребывание эмбрионов и личинок в нерестовых буграх, сут | 270-300 | 290-320 |
| плодовитость самок (нечетное поколение) 2001 г. 2003 г. | 1537 | 1758 2018 |
*данные Н. В. Гордеевой, 2003 г.
В новом регионе обитания горбуша сохранила присущие ей в нативном ареале основные ха - рактеристики нерестового хода, такие как волнообразная динамика миграции, увеличение к концу хода доли, как самок, так и более крупных производителей и другие.
Установлено, что к началу августа в море горбуша практически полностью перестает питаться, так, что относительная масса пищеварительного тракта с пищей снижается до 0,5%. В то же время относительная масса яичников последовательно увеличивалась и достигла к началу августа ¼ общей массы тела, что характерно для самок горбуши непосредственно перед нерестом. Дефинитивных размеров достигала и масса ооцитов. Таким образом, все исследованные производители потенциально готовы были нереститься до 15 сентября, т.е. в сроки, которые согласно проведенным ранее исследованиям (Дягилев, Маркевич, 1979), являются благоприятными для последующего выживания и развития молоди (Dorofeyeva et al., 2004).В настоящее время у всей исследованной молоди самок из реки Сояны, независимо от их длины и массы, к началу покатной миграции в яичниках уже сформирована генерация сходных по размеру ооцитов периода превителлогенеза. Таким образом, состояние гонад у самок в период покат- ной миграции не отличается от состояния гонад у молоди горбуши из рек Сахалина и Итурупа (Зе- ленников, Федоров, 2005). Следовательно, у нечетной линии беломорской горбуши, не происходит
замедления темпа раннего гаметогенеза, и нерест этих самок будет происходить в более ранние сроки (Дорофеева и др., 2007). Из этого следует один важный вывод - на протяжении 10 поколений (после последнего завоза икры нечетной линии из Магаданской области) в результате естественного отбора в популяции беломорской горбуши сохранились, в основном, ранненерестующие особи, т.е. произошла саморегуляция.
Сравнительные исследования пластических признаков беломорской популяции четной и нечетной линий демонстрируют сдвиги в ее морфологическом облике. Еще на первом этапе акклиматизации было отмечено (Суркова, 1970), что уже через 3-5 поколений беломорская горбуша стала иметь меньше позвонков и чешуй в боковой линии, у нее уменьшилась голова и соответственно ее пропорции. Результаты анализа 17 пластических признаков четной и нечетной популяций, проведенного на последнем этапе акклиматизации (2001-2003 гг.), также свидетельствуют о существенных сдвигах в их морфологическом облике по сравнению с рыбами из нативного ареала, причем сдвиги более выражены у четной линии и, по-видимому, инадаптивны (как результат катастрофического отбора), в то время как нечетная линия на фоне сравнительно высокой численности демонстрирует процесс адаптации к новым условиям (Гордеева, 2003).
При этом необходимо учитывать, что некоторая доля изменений морфологических признаков связана с деятельностью рыбоводных заводов и с гибридизацией между вновь вселенной магаданской популяцией и несомненно существующего небольшого числа производителей от предыдущего вселения горбуши из рек Сахалина. Пластические и некоторые меристические признаки беломорской горбуши сахалинского и магаданского происхождения имеют ряд существенные различий, и в настоящее время не представляется возможным определить степень влияния, оказанного потомками первой волны интродукции на формирование облика современного стада, с одной стороны, и, с другой стороны, изменения его под действием новых условий существования (Табл. 2).Таблица 2
Сравнительная характеристика морфологических признаков беломорской горбуши «сахалинского» и «магаданского» происхождения
| Признак | *»Сахалинская» 1965г. р. Сидоровка, n=50 | «Магаданская» 2005г. р. Кереть, n=65 |
| Длина тела (см) | 36-51 | 39-52 |
| Чешуй в LL | 156-185 (169) | 150-176 (169) |
| В % длины тела | ||
| Антедорсальное расстояние | 39-49 (44,4) | 41-48 (46,4) |
| Антеанальное расстояние | 61-68 (63,4) | 65-73 (61,2) |
| Высота D | 8-12 (9,1) | 10-13 (11,7) |
| Высота А | 8-10 (7,8) | 9-11 (10,5) |
| В % длины головы | ||
| Длина рыла | 24-38 (29,1) | 24-30 (26,9) |
| Диаметр глаза | 7-16 (11,5) | 12-15 (14,1) |
* По данным Е . И . Сурковой (1970)
Остеологические исследования беломорской горбуши ранее не проводились. Остеологическая изменчивость между локальными стадами у тихоокеанских лососей выражена сравнительно слабо, четкие клины отсутствуют, но определенные тенденции все-таки существуют (Горшков, 1979). Особенно информативными являются остеологические признаки, на которых в значительной мере основана классификация дальневосточных лососей: сошник, язычная кость и супраэтмоидеум.
Vomer (Рис. 2А). Форма сошника служит главным остеологическим признаком в систематике лососевых. Головка сошника сравнительно широкая, чаще треугольной формы, передняя ее часть вытянута, рукоятка лодкообразная, боковые края пластинчатые. При сравнении сошников горбуши из разных локальных стад можно отметить изменчивость формы головки и рукоятки. У беломорской горбуши сошники отличаются сравнительно широкой головкой и рукояткой, плавно сужающейся каудально. Эти признаки свойственны горбуше из реки Большой (Камчатка). Наибольшие отличия по форме головки у горбуши из реки Фирсовка (Сахалин).
Linguale (Рис. 2Б). Язычная кость у горбуши сравнительно узкая, наиболее широкая часть ее находится позади зубов, ряды зубов сходятся кпереди. По форме язычная кость беломорской горбуши наиболее сходна с язычной костью горбуши из рек Камчатки, а наибольшие отличия имеются у сахалинской горбуши.
Supraethmoideum (Рис. 2В). Супраэтмоидеум относительно широкая кость, в передней части
слега уплотненная, боковые лопасти тонкие, складки на них выражены слабо. На переднем конце
кости имеется небольшая головка. Относительная глубина вырезки по заднему краю не более 1520% общей длины кости. По форме этой кости беломорская горбуша сходна с горбушей из рек Камчатки, а по глубине вырезки с горбушей из реки Анадырь. Наибольшие отличия отмечены у горбуши из реки Фирсовка (Сахалин).
Таким образом, у сформировавшейся в Белом море популяции горбуши проявляются признаки, свойственные другим, как более северным (Анадырь), так и наиболее близким географическим популяциям (Камчатка).

Рис. 2. Морфологические ряды костей черепа Oncorhynchus gorbuscha из разных регионов ареала (рисунки и фотографии):
А - vomer: 1, 1a - р. Кереть; 2 - река Умба; 3, 3а - р. Большая (Камчатка); 4, 4а - р. Фирсовка (Сахалин); Б - Iinguale; обозначения те же; В - Supraethmoidale: 1, 1а - р. Кереть (2003 г.); 2 - р. Умба (2004 г.); 3 - р. Кереть (2005 г.); 4, 4а - р. Анадырь; 5, 5а - р. Большая (Камчатка); 6, 6а - р. Фирсовка (Сахалин).
Обращает на себя внимание факт, что у исследованных рыб из разных беломорских рек четной и нечетной линии остеологические особенности очень сходны и в равной мере отличаются от особенностей тихоокеанских стад, что вероятно связано с ограниченным числом донорских популяций.
Большой вклад в познание формирования популяции беломорской горбуши внесли исследования, проведенные в институте Общей генетики РАН (Гордеева, 2003; Гордеева, Салменкова и др., 2003). Выявлены существенные генетические изменения по сравнению донорской популяцией (редукции гетерозиготности и аллельного разнообразия и разнообразия гаплотипов мтДНК). Оказалось, что уровень генетических изменений существенно выше в нечетной линии, чем в четной, что на фоне успешного естественного воспроизводства у первой свидетельствует о процессе адаптации. «Четная линия» горбуши значительно больше специализирована на генетическом уровне, что, возможно, является причиной ее малой эффективности при акклиматизации.
На основании проведенных исследований, можно сделать следующие выводы. Нечетная линия горбуши, акклиматизированная в Белом море на протяжении 10 поколений, сохраняет устойчивые подходы производителей, что может свидетельствовать о формировании ранненерестующей популяции. Нечетная линия горбуши обнаруживает существенные морфобиологические и генетические сдвиги, которые на фоне устойчивых уловов позволяют судить о протекающих процессах адаптаций формирующейся беломорской популяции горбуши. Формирование ранненерестующей популяции реализуется, в основном, за счет адаптаций в процессе полового созревания (у покатной молоди отсутствует замедление темпа раннего онтогенеза, идущие на нерест самки готовы отнереститься в благоприятные для развития молоди сроки).
Работа поддержана грантом «Биологические ресурсы России» -II.2.4.
Литература
А.П. Алексеев, Е.А. Дорофеева, О.В. Зеленников. 2005. Современные представления о результатах интродукции дальневосточной горбуши в бассейн Белого моря // Материалы Международной специализированной конференции «Акватерра - 2005». Санкт-Петербург, 14-16 июня 2005 г., «Рестэк», СПб. [420 с.] С. 261264.
А.П. Алексеев, Е.А. Дорофеева, О.В. Зеленников. 2005. Современные представления о результатах интродукции дальневосточной горбуши в бассейн Белого моря // VIII Международная конференция и выставка «Акватерра - 2005». С. Петербург, ГУП «Водоканал». С. 261-264.
Алексеев А.П., Дорофеева Е.А., Зеленников О.В. 2008. Проблемы и перспективы акклиматизации дальневосточной горбуши в бассейне Белого моря // Рыбоводство и рыбное хозяйство. № 8. С. 24-28.
Алтухов Ю.П. 1974. Популяционная генетика. М. 247 с.
Воловик С.П. 1968. О возможности применения метода морфометрии для определения локальных популяций горбуши // Изв. Тихоокеанск. научн.-исслед. ин-та рыбн. хоз-ва и океанограф. Т. 65. С. 97-107.
Гордеева Н.В. 2003. Генетические процессы у горбуши интродуцированной в бассейне Белого моря // Автореферат к.б.н. 24 с.
Гордеева Н.В., Салменкова Е.А., Алтухов Ю.П., Махров А.А., Пустовойт С.П. 2003. Гордеева Н.В., Салменкова Е.А., Алтухов Ю.П., Махров А.А., Пустовойт С.П. 2003. Генетические изменения у горбуши Oncorhynchus gorbuscha (Walbaum) в ходе акклиматизации в бассейне Белого моря // Генетика. Т. 39, № 3. С. 402-412.
Горшков С.А. 1979. Сравнительно-морфологические основы классификации тихоокеанских лососей рода Oncorhynchus. Кандидатская диссертация. ЗИН АН СССР. 260 с.
Горшков С.А., Дорофеева Е.А., Клюканов В.А., Куликова Н.И. 1979. Остеологические особенности тихоокеанских лососей рода Oncorhynchus // Вопр. ихтиол. Т. 19, вып. 6 (119). С. 963-982.
Е.А. Дорофеева, А.П. Алексеев, В.Г. Кулачкова, О.В. Зеленников, Т.С. Иванова. 2004. Актуальные проблемы акклиматизации горбуши в Белом море // В материалах конференции: «Проблемы изучения, рационального использования и охраны ресурсов Белого моря». Петрозаводск. Институт биологии Карельского НЦ РАН. С. 105-109.
Дорофеева Е.А., Алексеев А.П., Зеленников О.В. «Морфологические особенности формирования популяций дальневосточной горбуши в Белом море». Материалы 2-й международной конференции «Экологические исследования беломорских организмов». Июнь 2007 г., м. Картеш. С. 34-36.
Дорофеева Е.А., Зеленников О.В., Боркичев В.С., Алексеев А.П. «Формирование популяций горбуши в белом море». Материалы X Международной конференции «Проблемы изучения, рационального использования и охраны природных ресурсов Белого моря». Сентябрь 2007 г. Архангельск. С. 116-122.
Дорофеева Е.А., Алексеев А.П., Зеленников О.В., Зеленков В.А. Дальневосточная горбуша в бассейне Белого моря (к пятидесятилетию интродукции) // Рыбное хозяйство. № 6. 2006. С. 71-73.
Алексеев А.П., Дорофеева Е.А., Зеленников О.В. 2008. Проблемы и перспективы акклиматизации дальневосточной горбуши в бассейне Белого моря // Рыбоводство и рыбное хозяйство. № 8. С.24-28.
Дягилев С.Е., Маркевич Н.Б. 1979. Разновременность созревания горбуши Oncorhynchus gorbuscha (Walb.) четных и нечетных лет как основной фактор, определивший различные результаты ее акклиматизации на севере Европейской части СССР // Вопр. ихтиол. Т. 19, вып. 2. С. 230-245.
Енютина Р. И. 1972. Амурская горбуша (промыслово-биологический очерк) // Изв. Тихоокеанск. на- учн.-исслед. ин-та рыбн. хоз-ва и океанограф. Т. 77. С. 3-126.
Персов Г.М. 1975. Дифференцировка пола у рыб. Л.: Изд-во ЛГУ. 148 с.
Пахомова Н.А., Хлевная А.С. 1978. Морфология яичников горбуши в морской период жизни // Вопросы функциональной морфологии морских животных. Апатиты. С. 102-108.
Персов Г.М., Федоров К.Е., Сакун О.Ф., Чистова М.Н. 1983. Биологические основы, биотехника и надежность процесса акклиматизации дальневосточной горбуши Oncorhynchus gorbuscha (Walbaum) (Salmonidae) на Европейском Севере СССР // Вопросы ихтиол. Т. 23, вып.4. С.622-628.
Сакун О.Ф. 1965. Возникновение изменений в гаметогенезе и половом цикле у горбуши Oncorhynchus gorbuscha (Walb.) за пределами естественного ареала // Вопросы ихтиол. Т. 5, вып. 4. С. 639-651.
Суркова Е. И. 1970. Изменение морфологических признаков горбуши в ходе ее акклиматизации на Европейском Севере // Материалы рыбохозяйственных исследований Северного бассейна. Вып.14. С.78-85.
Dorofeyeva Ye.A., A.P. Alekseev, O.V. Zelennikov, T.S. Ivanova. 2004. Acclimatization of Far Eastern pink
salmon in the White Sea: results and perspectives // Proc. in Zool. Inst. RAS. V. 300. P.53-63.
Hoard W.R. 1991. Life history of pink salmon (Oncorhynchus gorbusha). Pacific Salmon Life Histories (Eds Groot C., Margolis L.). Vancouver: UBC Press. P. 119-230.
Lear W.H. 1975. Evaluation of transplant of Pacific salmon (Oncorhynchus gorbuscha) from British Columbia to Newfoundland // О. Fish. Res. Board Canada. V. 32, No 12. P. 2343-2356.
MORPHOLOGICAL ADAPTATIONS OF PINK SALMON IN THE WHITE SEA E.A. Dorofeyeva
Zoological Institute, Russian Academy of Sciences, Saint-Petersburg, Russia e-mail: salmo@zin.ru
As a result of studies of White Sea pink salmon carried out in 2002-2007 has been concluded that at present the odd-year line of pink salmon after the last introduction has retain for 11 generations stable spawning runs and reveals adaptation directed to for formation of early spawning population. The rate of development of sex dlands of fish in earlier age and further in course of sea migration allows the entire generation of pink salmon to attain sexual maturity in period favourable for subsequent development of the young. The odd-line generation had demonstrated the change of morphological and genetic characters (the plastic, meristic and osteological characters, the lower value of heterozygosity, allele diversity and diversity of haplotypes of mt DNA) suggest adaptation process.
Еще по теме МОРФОБИОЛОГИЧЕСКИЕ АДАПТАЦИИ ГОРБУШИ В БЕЛОМ МОРЕ:
- ЭКОЛОГИЯ ДЕТЕНЫШЕЙ ГРЕНЛАНДСКОГО ТЮЛЕНЯ НА РАЗНЫХ СТАДИЯХ РАЗВИТИЯ В БЕЛОМ МОРЕ
- ПИТАНИЕ METRIDIA LONGA (COPEPODA; CALANOIDA) В БЕЛОМ МОРЕ
- МОНИТОРИНГ МОРСКИХ МЛЕКОПИТАЮЩИХ В БЕЛОМ МОРЕ ЛЕТОМ 2007-2008 гг.
- ПРОБЛЕМЫ РЫБОЛОВСТВА В БЕЛОМ МОРЕ И ПУТИ ИХ РЕШЕНИЯ А.С. Шерстков, С.Б. Фролов, В.С. Шерстков
- 1.1.6. Опросник «Шкала семейной адаптации и сплоченности» (FACES - III) Д.Олсона в адаптации А.Г. Лидерса и Е.Н. Спиревой
- 1.2.3. Понятие и сущность адаптации. Социально-психологическая адаптация.
- 25 и 26. Адаптация персонала. Цели и этапы адаптации
- Повесть о белом клобуке .
- «О ТВЕРДОМ И БЕЛОМ»
- НОВЫЙ ЧЕЛОВЕК В БЕЛОМ ДОМЕ