ПРОДУКЦИОННЫЕ ХАРАКТЕРИСТИКИ ФИТОПЛАНКТОНА БЕЛОГО МОРЯ В ЗАВИСИМОСТИ ОТ ИСТОЧНИКА АЗОТА
Т.А. Белевич
Московский государственный университет им. М.В.Ломоносова, г. Москва, Россия e-mail: 3438083@list.ru
В природных экосистемах обеспеченность фитопланктона минеральными и органическими ресурсами, а также световые условия меняются как во времени, так и в пространстве.
В Белом море, как и в других морях умеренных и высоких широт весеннее развитие фитопланктона ведет к практически полному исчерпанию нитратного азота в фотическом слое ( Максимова, 1991; Ильяш и др., 2003). Последующая деструкция биомассы, накопленной при весеннем «цветении» и выделение органического азота зоопланктоном ведет к увеличению концентрации органического азота и аммония. Соответственно, доля восстановленных форм азота в суммарном содержании растворенного азота увеличивается (Максимова, 1991; Ильяш и др., 2003). При этом в поверхностном слое фитопланктон испытывает стресс фотоингибирования, на промежуточных глубинах фотической зоны освещенность близка к насыщающему фотосинтез уровню, на нижней границе фотической зоны освещенность лимитирует фотосинтез (Falkowski, Raven, 1997). В летний период при низкой концентрации минеральных ресурсов в ряде случаев наблюдается интенсивное развитие фитопланктона. Как это показано для Белого моря (Ильяш и др., 2003), в основном это миксотрофные водоросли. Увеличение биомассы фитопланктона при низкой обеспеченности нитратами связывают со способностью многих популяций водорослей использовать органические формы азота (Иль- яш и др., 2003; Antia et al., 1991; Berman, Bronk, 2003). Увеличение относительной доли органического азота в пуле растворенного азота ведет к изменению структуры фитопланктона (Berg et al., 2003; Запара и др., 2007). На примере фитопланктона Белого моря показано, что ассимиляция водорослями разных азот содержащих субстратов (мочевины, глицина, нитратов, аммония) вела к формированию различающихся по структуре сообществ (Запара и др., 2007).Ранее на смешанных культурах было выявлено, что динамика биомассы водорослей видоспецифично зависит не только от источника азота, но и от уровня освещенности (Ильяш, Запара, 2006). Последнее, с учетом различающихся энергетических и субстратных затрат водорослей при ассимиляции восстановленных и окисленной форм азота (Huppe, Turpin, 1994), дает основание ожидать, что продукционные характеристики и природного фитопланктона будут зависеть от источника азота и освещенности.
В настоящем исследовании проведен сравнительный анализ динамики биомассы и продукционных характеристик экспериментальных сообществ фитопланктона Белого моря при ассимиляции нитратов, аммония, глицина и мочевины при двух уровнях освещенности.
Материал и методы
Эксперименты проводили на Беломорской биологической станции Московского государственного университета (Карельский берег Кандалакшского залива Белого моря) с 23 августа по 10 сентября 2007 г. В позднелетний период фитопланктон Белого моря лимитирован недостатком азота (Максимова, 1991; Ильяш и др., 2003). Согласно многолетним наблюдениям, концентрация минерального азота в Кандалакшском заливе в этот период не превышает 3 мкмоль/л (Максимова, 1991).
Схема эксперимента. Фитопланктон, служивший исходным материалом для экспериментов, отбирали с помощью сети из планктонного газа № 78 в слое 2-5 м. Для устранения пресса выедания растительноядным зоопланктоном фитопланктон пропускали через планктонный газ № 40. В 1,5 литровые пластиковые ёмкости добавляли отфильтрованную морскую воду, концентрированный фитопланктон (посевной титр - 1150 кл/мл, 2940 мкгС/л), а так же все биогенные элементы, за исключением азота, согласно прописи среды f/2 (Guillard, Ryther, 1962). Азот вносили в виде мочевины, глицина, нитратов или аммония в концентрации 180 мкмоль азота/л. Соотношение содержания азота и фосфора в среде равнялось пяти, что согласно общепринятым взглядам (Ryther, Dunstan, 1971), обусловливает ограничение развития водорослей недостатком азота.
В качестве контроля использовали фитопланктон без добавок азота. Экспериментальныеемкости экспонировали in situ на плотиках на глубине 1 м. Полуденная освещенность на этой
глубине (E1) колебалась в пределах 25 - 1050 мкЕ/(м2 сек). Более низкую освещённость (E2), составлявшую в среднем 51 % от E1, создавали путём экранирования склянок тканью средней плотности.
Все варианты эксперимента проводили в трёх повторностях. При дальнейшем изложении сообщества, росшие с использованием разных источников азота, обозначены следующим образом: сообщество, ассимилирующее нитраты - N, глицин - G, мочевину - M, аммоний - A. Контрольное сообщество (без добавок азота) обозначено как K.
Оцениваемые параметры. Раз в трое суток из каждой емкости проводили отбор проб объёмом 20 мл и фиксировали их раствором Люголя. Отобранные пробы, представляющие повторности для каждого варианта добавки и контроля (по три пробы), сливали и микроскопическому анализу подвергали интегральную пробу. Численность водорослей (Nn) определяли методом прямого счета в камере Нажотта (объемом 0.05 мл). Число просчитанных клеток превышало 3000. При этом ожидаемая относительная ошибка измерений оказывается в пределах 5-10% (Федоров, 1979). По методу геометрического подобия определяли объемы клеток (Hillebrand et al., 1999). Для оценки величин биомассы в единицах углерода клеточное содержание органического углерода в зависимости от объема клеток рассчитывали по аллометрическим уравнениям (Menden-Deuer, Lessard, 2000). Фотосинтетическое выделение кислорода оценивали скляночным методом в кислородной модификации (Винберг, 1960). Продукционные склянки инкубировали на плотиках на глубине 1 м. Склянки с фитопланктоном, росшим при освещенности E2, экранировали тканью средней плотности. Время экспозиции составляло 4 ч, объем экспонируемых проб - 30 мл. Концентрацию кислорода измеряли с помощью оксимера Oxi 330. По величинам фотосинтетического выделения кислорода согласно балансовому уравнению фотосинтеза (Винберг, 1960) рассчитывали фотосинтетическую фиксацию углерода.
Результаты и обсуждение
В составе экспериментальных сообществ фитопланктона отмечено 55 таксономических единиц, относящихся к Bacillariophyceae и Dinophyta. Диатомовые водоросли составили 87% от общего числа видов. В исходном сообществе доминировала водоросль Ditylum brightwelii. Ее вклад в суммарную численность составил 32%, а вклад в суммарную биомассу достигал 76%.
В контрольных сообществах (без добавок азота) биомасса фитопланктона увеличивалась до 6 сут эксперимента (рис.). Рост водорослей, по-видимому, происходил за счет внутриклеточных запасов азота. Доминирование D.brightwelii сохранялось на протяжении всего эксперимента. На 6 сут, когда суммарная биомасса в контрольных сообществах достигала максимума, вклад D.brightwelii в суммарную биомассу составлял 96% при обеих освещенностях. Величина накопленной биомассы (Bmax) была выше при более низкой освещенности (рис.). Это свидетельствует о том, что при E1 лимитированные по азоту водоросли, и прежде всего доминирующая в сообществе водоросль D.brightwelii, испытывали стресс фотоингибирования. Это также подтверждается более низкими значениями удельной фотосинтетической фиксации углерода (Р/В-коэффициента) контрольных сообществ при E1, чем при E2. (табл.). При воздействии ингибирующей интенсивности света часть клеточного вещества и энергии водорослей затрачивается на восстановление фотоиндуцируемых повреждений фотосинтетического аппарата (Falkowski, Raven, 1997), а также на более интенсивный синтез светозащитных пигментов (Ben-Amotz et al., 1989), что снижает количество вещества и энергии, направляемых на рост. При увеличении биомассы водорослей в контрольных сообществах степень их лимитирования недостатком азота возрастает. По мере увеличения степени азотного дефицита у водорослей чувствительность к фотоингибированию повышается (Kolber et al., 1988), а способность к фотоакклимации снижается (Herzig, Falkowski, 1989).
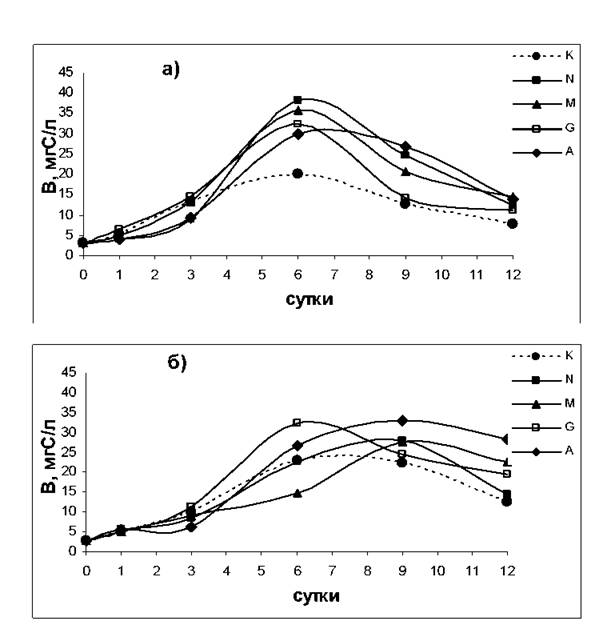
Динамика биомассы фитопланктона (B) в присутствие добавок нитратов (N), глицина (G), мочевины (М), аммония (А) и без добавок (К) при освещенности E1 (а) и E2 (б).
Динамика удельной фотосинтетической фиксации углерода (Р/B, 10-2 сут-1) у водорослей, росших с добавками нитратов (N), мочевины (М), глицина (G), аммония (А) и без добавок азота (К) при освещенности Е1 и Е2.
| Сутки роста | Освещенность | |||||||||
| E1 | E2 | |||||||||
| Добавки | Добавки | |||||||||
| K | N | M | G | A | K | N | M | G | A | |
| 1 | 9,3 | 4,5 | 11,9 | 11,9 | 4,7 | 13,1 | 8,2 | 10,3 | 13,6 | 10,1 |
| 3 | 2,2 | 12,5 | 6,1 | 5,5 | 7,0 | 9,2 | 8,0 | 9,5 | 7,9 | 6,0 |
| 6 | 4,2 | 5,1 | 6,7 | 9,5 | 6,6 | 6,4 | 7,6 | 11,6 | 9,9 | 2,9 |
| 9 | 4,3 | 4,2 | 1,3 | 3,5 | 5,8 | 5,0 | 6,1 | 7,9 | 10,4 | 5,2 |
После внесения добавок азота во всех сообществах наблюдалось увеличение суммарной биомассы водорослей, превосходящее таковое в контроле (рис.). Величина накопленной биомассы фитопланктона зависела от источника азота и уровня освещенности.
В сообществах N и M, величинаBmax была выше при E1, в сообществе A - при E2, а сообществе G величины Bmax существенно не различались. Это свидетельствует о том, что для водорослей, обеспеченных азотом нитратов и мочевины, освещенность Е2 являлась лимитирующей. При ассимиляции нитратов на их восстановление используется часть восстановителя и АТФ, образованных в световых реакциях фотосинтеза (Falkowski, Raven, 1997). Потребление мочевины идет за счет активного транспорта (Williams, Hodson, 1977) с использованием в качестве основного источника энергии фотогенерированной АТФ (Rees, Syrett, 1979). Потребность при ассимиляции нитратов и мочевины в продуктах световых реакций фотосинтеза и обусловило более высокие значения Bmax при лучшей обеспеченности световой энергией. Величина Bmax сообщества N была выше, чем в сообществе М. По-видимому, суммарные энергетические затраты водорослей на активный транспорт и последующую внутриклеточную трансформацию мочевины оказались выше затрат на восстановление азота нитратов. Хотя отмечено, что некоторые виды водорослей при использовании в качестве источника азота мочевины растут с большей скоростью, чем при использовании нитратов (Antia et al., 1991).
При акклимации к лимитирующей освещенности эффективность фотосинтетической фиксации углерода уменьшается, и фотосинтез на единицу хлорофилла (или на единицу компонента электронного транспорта) снижается (Sukenik et al., 1990; Falkowski, Raven, 1997). Снижение удельной фотосинтетической фиксации углерода может быть компенсировано за счет углерода, входящего в состав ассимилируемых органических субстратов и, в частности, глицина и мочевины (Antia et al., 1991; Mulholland et al., 2003). Преимущество покрытия потребностей не только в азоте, но и в углероде при лимитирующей освещенности за счет использования одного субстрата не проявилось ни при использовании глицина (величины Bmax при E1 и E2 существенно не различались), ни при росте с мочевиной (Bmax выше при E1).
При лимитирующей освещенности можно ожидать более эффективный рост фитопланктона с использованием восстановленного азота аммония по сравнению с ростом за счет потребления нитратов. При ассимиляции нитратов на их восстановление используется часть восстановителя и АТФ, образованных в световых реакциях фотосинтеза (Falkowski, Raven, 1997). Это ведет к уменьшению количества восстановителя и энергии, доступных для использования при фиксации углерода, что обусловливает меньшую скорость роста водорослей при ассимиляции нитратов, по сравнению с таковой при использовании восстановленного азота аммония (Huppe, Turpin, 1994). Преимущество роста с использованием восстановленного аммонийного азота при лимитирующей освещенности обусловило более высокую биомассу, накопленную сообществом А по сравнению с Bmax в сообществе N.
Динамика удельной фотосинтетической активности водорослей зависела от формы азота и освещенности (табл. 1). В первые сутки роста, когда идет интенсивное потребление добавок, наибольшие величины P/B отмечены при El в сообществах M и G, а при E2 - в сообществе G. На третьи сутки, когда потребление азота, по-видимому, закончилось (в силу небольшой концентрации добавок), наибольшие значения P/B достигались в сообществах N при El и М при E2. В период достижения Bmax (6 сут при El и 9 сут при E2) наибольшие значения P/B отмечались в сообществах G. Сопоставление величин удельной фотосинтетической активности водорослей, ассимилирующих ту или иную добавку при двух уровнях освещенности (табл. l), показывает, что в период активного роста P/B выше при E2, чем при El в сообществах, росших с использованием глицина, мочевины (кроме первых сут) и нитратов (кроме третьих суток). Напротив, в сообществах, росших с использованием аммония P/B выше при El, чем при E2 за исключением первых суток.
Таким образом, результаты обогатительных экспериментов показали, что динамика биомассы и продукционных характеристик фитопланктона зависит от источника азота и освещенности. Это, по-видимому, обусловлено различающимися энергетическими затратами на ассимиляцию восстановленных и окисленной формы азота, а также тем, что разные формы азота используются отдельными водорослями на рост и другие метаболитные нужды с неодинаковой эффективностью (Fan et al., 2003), причем эффективность зависит от освещенности (Levasseur et al., l993).
Работа выполнена при финансовой поддержке Российского фонда фундаментальных исследований (грант № 08-04-00932).
Литература
Винберг Г.Г., l960. Первичная продукция водоемов. Минск: Изд-во АН БССР. 329 с.
Запара Е.В., Белевич Т.А., Ильяш Л.В., 2007. Конкурентные отношения между планктонными водорос
лями Белого моря при разных источниках азота // Журн. общ. биол. Т. 68, № 3. С. l95-204.
Ильяш Л.В., Запара Е.В., 2006. Конкуренция двух морских диатомовых водорослей за азот мочевины и
нитратов при трех уровнях освещенности // Журн. общ. биол. Т. 67, № 6. С. 464-475.
Ильяш Л.В., Житина Л.С., Федоров В.Д., 2003. Фитопланктон Белого моря. М.: Янус-К. l68 c. Максимова М.П., l99l. Гидрохимия Белого моря // Гидрометеорология и гидрохимия морей СССР. Т.2.
Белое море. Ч.1. С. 8-193.
Федоров В.Д., l979. О методах изучения фитопланктона и его активности. М.: Изд. МГУ. l65 с.
Antia N.J., Harrison J.P., Oliveira L., 1991. The role of dissolved organic nitrogen in phytoplankton nutrition,
cell biology and ecology // Phycologia. V. 30. P. 1-89.
Ben-Amotz A., Shaish V., Avron M., 1989. Mode of action of the massively accumulated β-carotene of Dunaliella bardawil in protecting the alga against damage by excess irradiation // Plant Physiol. V. 91. P. 1040-1043.
Berg G.M., Balode M., Purina I., Bekere S., Bechemin C., Maestrini S.Y., 2003. Plankton community composition in relation to availability and uptake of oxidized and reduced nitrogen //Aquat. Microb. Ecol. V. 30. P. 263-274.
Berman T, Bronk D.A., 2003. Dissolved organic nitrogen: a dynamic participant in aquatic ecosystems // Aquat. Microb. Ecol. V. 31. P. 279-305.
Falkowski P.G., Raven J.A., 1997. Aquatic photosynthesis. Malden: Blackwell Science. 375 p.
Fan C., Glibert P.M., Lomas M.W., 2003. Characterization of urease activity in three marine phytoplankton species, Aureococcus anophagerefferens, Prorocentrum minimum, and Thalassiosira weissflogii // Mar. Biol. V. 142.
P. 949-958.
Guillard R. R. L., Ryther J. H., 1962. Studies on marine diatoms. I. Cyclotella nana Hustedt and Detonula confervacea (Cleve) Gran. // Can. J. Microbiol. № 8. P. 229-239.
Herzig R., Falkowski P.G., 1989. Nitrogen limitation in Isochrysis galbana (Haptophyceae). Photosynthetic energy conversion and growth efficiencies // J. Phycol. V. 25. P. 462-471.
Hillebrand H., Durselen C. D., Kirschtel D., Pollingher U., Zohary T., 1999. Biovolume calculation for pelagic and benthic microalgae // J. Phycol. V. 35. P. 403-424.
Huppe H.C., Turpin D.H., 1994. Integration of carbon and nitrogen metabolism in plant and algal cells // Ann. Rev. Plant Physiol. Plant Mol. Biol. V. 45. № 3. P. 577-607.
Kolber Z., Zehr J., Falkowski P.G., 1988. Effects of growth irradiance and nitrogen limitation on photosynthetic energy conversion in Photosystem II // Plant Physiol. V. 88. P. 923-929.
Levasseue M., Thompson P.A., Harrison P.J., 1993. Physiological acclimation of marine phytoplankton to different nitrogen sources // J. Phycol. V. 29. P. 587-595.
Menden-Deuer S., Lessard D.J., 2000. Carbon to volume relationships for dinoflagellates, diatoms, and other protist plankton // Limnol. Oceanogr. V. 45. P. 569-579.
Mulholland M.R., Lee C., Glibert P.M., 2003. Extracellular enzyme activity and uptake of carbon and nitrogen along an estuarine salinity and nutrient gradient // Mar. Ecol. Prog. Ser. V. 258. P. 3-17.
Rees T.A.V., Syrett P.J., 1979. Mechanisms for urea uptake by the diatom Phaeodactylum tricornutum: the uptake of thiourea // New Phytol. V. 83. P. 37-48.
Ryther J., Dunstan W.M., 1971. Nitrogen, phosphorus and eutrophication in the coastal marine environment // Science. V. 171. P. 1008-1013.
Sukenik A., Bennett J., Mortain-Bertrand A., Falkowski P.G., 1990. Adaptation of the photosynthetic apparatus to irradiance in Dunaliella tertiolecta // Plant Physiol. V. 92. P. 891-898.
Williams S.K., Hodson R.C., 1977. Transport of urea at low concentrations in Chlamydomonas reinhardtii // J. Bacteriol. V. 130. P. 266-273.
THE PRODUCTIONAL CHARACTIRISTICS OF PHYTOPLANKTON OF THE WHITE SEA IN DIFFERENT SOURCE OF NITRATE Т.А. Belevich
Moscow State University, Moscow, Russia e-mail: 3438083@list.ru
Phytoplankton was sampled in the Kandalaksha Bay of the White Sea in the end of august, 2007. The samples were kept 14 days in situ under two levels of irradiance (Е1>Е2) with additions of nitrogen in form of urea (U), glycine (G), nitrate (N) and ammonium (A). Species composition, abundance, biomass and primary production were estimated every 3rd day. After additions of nitrogen increase of gross biomass was registered in all experimental communities. This increase was considerably higher then in control. Biomass gain of phytoplankton depended on nitrogen source and irradiance level. In N and U communities Bmax was higher under E1, in A community - under E2, in G community Bmax didn't snow any considerable differences. The comparison of specific photosynthetic activity of algae, assimilating one or another addition of nitrogen under two levels of irradiance, shows, that during a period of active growth in G and U communities (except 1st day) and in N community (except 3rd day) P/B was higher under E2, than under E1. At the same time, in A communities P/B was higher under E1, than under E2 (except 1st day).
Еще по теме ПРОДУКЦИОННЫЕ ХАРАКТЕРИСТИКИ ФИТОПЛАНКТОНА БЕЛОГО МОРЯ В ЗАВИСИМОСТИ ОТ ИСТОЧНИКА АЗОТА:
- О ЛЕТНЕМ ИХТИОПЛАНКТОНЕ ДВИНСКОГО ЗАЛИВА БЕЛОГО МОРЯ
- ПЕРВИЧНАЯ ПРОДУКТИВНОСТЬ ФИТОПЛАНКТОНА В ПРОЛИВЕ ПЕЧАКОВСКАЯ САЛМА БЕЛОГО МОРЯ В ЛЕТНИЙ ПЕРИОД.
- ВЛИЯНИЕ РАЗНОЙ СОЛЕНОСТИ НА ЛИПИДЫ АМФИПОД БЕЛОГО МОРЯ
- СОСТАВ КРИОФЛОРЫ ПРИБРЕЖНЫХ ЛЬДОВ КАНДАЛАКШСКОГО ЗАЛИВА БЕЛОГО МОРЯ
- СОСТОЯНИЕ ГИДРОБИОЦЕНОЗОВ РЕК ПОМОРСКОГО ПОБЕРЕЖЬЯ БЕЛОГО МОРЯ
- СЕЗОННО-ОБУСЛОВЛЕННЫЕ ИЗМЕНЕНИЯ В ТРАНСМИССИИ ПАРАЗИТОВ В ПРИБРЕЖЬЕ БЕЛОГО МОРЯ
- КРИОФАУНА ЛЬДОВ ПРОЛИВА ВЕЛИКАЯ САЛМА КАНДАЛАКШСКОГО ЗАЛИВА БЕЛОГО МОРЯ
- СТРУКТУРА И ДИНАМИКА ПРИМОРСКИХ РАСТИТЕЛЬНЫХ СООБЩЕСТВ ПОМОРСКОГО И КАРЕЛЬСКОГО БЕРЕГОВ БЕЛОГО МОРЯ Л.А. Сергиенко
- ДИНАМИКА СТРУКТУРЫ ПОСЕЛЕНИЯ MYTILUS EDULIS НА ЛИТОРАЛИ КАНДАЛАКШСКОГО ЗАЛИВА БЕЛОГО МОРЯ
- РАЗМЕРНЫЙ СОСТАВ И ЛИПИДНЫЕ ПОКАЗАТЕЛИ У АМФИПОД БЕЛОГО МОРЯ В РАЗНЫХ УСЛОВИЯХ ОБИТАНИЯ Г.А. Шкляревич1, В.В. Богдан2
- К ЭКОЛОГИИ ПРИМОРСКИХ ОСОК (CAREX L) КАНДАЛАКШСКОГО ЗАЛИВА БЕЛОГО МОРЯ
- МОРФОЛОГИЧЕСКИЕ АСПЕКТЫ ВИДООБРАЗОВАНИЯ И ФОРМИРОВАНИЯ АДАПТАЦИОННЫХ СТРАТЕГИЙ ВИДОВ Р. COCHLEARIA L. В ПРИБРЕЖНЫХ РАСТИТЕЛЬНЫХ СООБЩЕСТВАХ БЕЛОГО МОРЯ Л.А. Сергиенко
- ПОЛОВАЯ СТРУКТУРА МОРСКОЙ ЗВЕЗДЫ ASTERIAS RUBENS L. НА ТЕРРИТОРИИ КАНДАЛАКШСКОГО ЗАЛИВА БЕЛОГО МОРЯ В 2003 ГОДУ
- НОВЫЕ ДАННЫЕ О ВОДНЫХ БЕСПОЗВОНОЧНЫХ БЕЛОГО МОРЯ В ПИТАНИИ ЛЕСНЫХ ПТИЦ