КЛАСС ASPIDOGASTRAEA FAUST ЕТ TANG
Аспидогастриды представляют собой небольшую группу паразитических плоских червей, до недавнего времени относившуюся к классу трематод. С последними их сближают многие черты внутренней организации, в сущности характерные для всего типа Plathelminlhes.
Объединение же обеих групп в один класс невозможно из-за принципиальных различий в ходе их жизненного цикла.Аспидогастриды известны с 1827 г., когда академик К. Бэр впервые опубликовал сведения о паразитах, найденных им в перикардиальной полости двухстворчатых моллюсков Лпосіопіа и Unio. Этот вид—Aspidogaster conchicola Baer, 1827,— считается типичным представителем класса. В дальнейшем многие исследователи занимались изучением аснндогастрид. Их место в системе было определено Б. Фаустом и К. Тангом (Faust et Tang), которые возвели эту группу в равный таксономический ранг с днгспстпчсскпмн и моногенетическнми сосальщиками. Разделение последних на два самостоятельных класса (Быховскпй, 1957) сделало неизбежным и обоснование самостоятельного класса Aspidogastraea. Жизненный цикл /1. conchicola был подробно исследован еще в конце прошлого века (Voll/kuw, 18УК). Строению аснндогастрид, их филогении и положенню в системе плоских червей посвящены работы Б. Е. Быковского, Г. Стэнкарда (Stunkard), К. Оденнш а (Odening), К. Роде (Rhode) и др. Все они в значительной мере дискуссионны.
Представители Aspidogastraea (рис. 145, А) паразитируют в моллюсках (в основном — пластинчатожаберных) и в водных позвоночных (рыбы и черепахи). По большей части это мелкие черви, 3—5 мм длиной. На переднем конце их тела располагается ротовое отверстие, окаймленное мускульным валиком. Оно ведет в неразветвленный мешковидный кишечник. Выделительная система аснндогастрид устроена по протопефрпдпальному типу; нервная — носит' характер ортогопа. Гермафродитная половая система (рис. 145, Б) мало чем отличается от таковой других плоских червей.
Самая примечательная особенность аспндогастрнд—наличие па брюшной стороне их тела большого прикрепительного диска (диск Бэра), отделенного от внутренних органов мышечной перегородкой, которая пересекает паренхиму.
Диск разбит на множество (50—70 и более) ячеек, каждая из которых окружена мускульным валиком и по сути дела может рассматриваться как небольшая самостоятельная присоска. Расположение и число этих присосок у разных видов аспндогастрнд различно. По наружному краю диска расположены многочисленные грушевидные образования — маргинальные (краевые) органы, до недавнего времени рассматривавшиеся как органы чувств. Однако исследования последних лет показали, что это не так. Диск Бэра несет многочисленные одноклеточные железы, часть которых продуцирует слизистый секрет, содержащий нейтральные мукополисахарнды. Этот секрет поступает по системе протоков в маргинальныеорганы и накапливается в полости их грушевидных вздутий, которые представляют собой не что иное, как своеобразные резервуары, специальным отверстием сообщающиеся с внешней средой. По-видимому, выделяемая наружу слизь играет известную роль в процессе прикрепления паразита, а возможно, способствует и его передвижению (скольжению) по телу хозяина. В этой связи обращает на себя внимание и характер иннервации прикрепительного диска. Здесь образуется нервное сплетение, состоящее из' системы нервных колец п связывающей их сети топких поверхностных волокон.
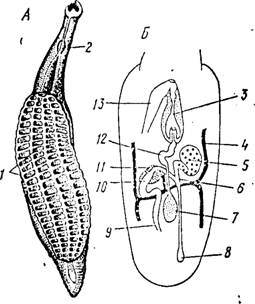
Рис. 145. Строение аснндогастрид. Л—Aspidogaster conchicola (по Стрелкову); Б— схема строения полового аппарата (на кп. Дауэса):
/ — диск Б>ра, 2 —- глотка, 3 — сумка цир* руса, V — желточный проток, 5 — яичник, 6 — яйцевод, 7 — семенник, 8 — лауреров канал, 9 — начало маткк, 10 — оотип, // — тельце Мелиса, 12 — семенной пузырек, 13 — концевой отдел матки
Судя но последним данным, диск Бэра участвует и процессах ініекпшечпого пищеварения. Тегумент ячеек диска обладает характерной ультраструктурой и песет многочисленные микровореннкп. В толще тегумеитн обнаружена высокая активность фосфатаз, здесь же открываются протоки одноклеточных белковых желез. Все это позволяет сравнивать диск Бэра с органом Брандеса трематод отр.
Strigeidida, который, как известно, выполняет функции внекишечного переваривания тканей хозяина и последующего всасывания продуктов гидролиза (с. 217).Аспидогастрндам свойствен простои жизненный цикл, проходящий без чередования поколений (рис. 146). У 'Некоторых видов (например, у Aspidogaster conchicola) эмбриональное развитие завершается еще во время пребывания яиц в матке червя. Наружу выносятся яйца, содержащие вполне сформированную, жизнеспособную личинку. Вылупленне последней происходит через несколько минут после попадания в воду. Личинка нс питается и живет, расходуя имеющийся в се тканях запас гликогена. В отличие от мирациднев трематод она лишена ресничек и передвигается за счет мускульных сокращений.
Заражение хозяина происходит пассивно. Личинки заносятся в мантийную полость моллюска вместе с током воды, поступающей через жаберный сифон.
Они мигрируют по телу хозяина, пока не достигнут своего местообитания. Некоторые виды локализуются непосредственно в мантийной полости, другие—в околосердечной сумке, в почке и различных внутренних органах. В процессе развития личинки ее брюшная присоска пое- образуется в сложно устроенный прикрепительный диск.
Аспидогастриды, паразитирующие у позвоночных (пресноводных и морских рыб и черепах), локализуются, как правило, в кишечнике. Жизненный цикл многих видов аспндогастрнд проходит со сменой животных хозяев. Промежуточным хозяином служат при этом моллюски, окончательным — позвоночные. Однако, экспериментальные данные показывают, что некоторые виды (Л. limacoides, например) способны завершать свое развитие в организме моллюска и их жизненный цикл может ограничиваться одним хозяином. В то же время такие виды, как A. conchicola, нормально паразитирующие только в моллюсках, будучи скормлены- рыбам, остаются живыми в их кишечнике на протяжении нескольких дней (Т. А. Тимофеева). На основании этих фактов можно предположить, что для многих видов аспидогастрид характерен простой жизненный цикл, связанный лишь с одним хозяином — моллюском. Позвоночные же, питаясь моллюсками, играют роль факультативных; случайных хозяев, в кишечнике которых паразиты остаются живыми в течение некоторого времени. Закрепление этой особенности
п филогенезе, по-шілнмому, и привело к возникновению видов, ставших специфичными паразитами позвоночных.
Таким образом, жизненные циклы аспидогастрид не имеют ничего общего с жизненными циклами трематод. Эти группы, хотя и связа'иы между собой общностью происхождения, несомненно должны быть отнесены к разным классам.
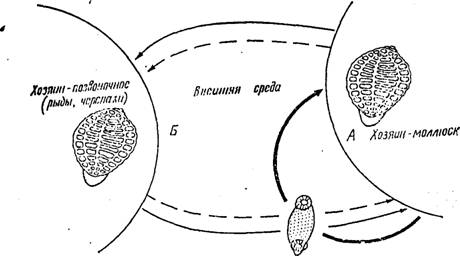
Рис. 146. Жизненный цикл аспидогастрид. А — цикл идет с участием одного хозяина; Б — цикл идет с участием двух хозяев (пунктиром показана факультативная возможность хода жизненного цикла у видов, нормально развивающихся по типу А)
Еще по теме КЛАСС ASPIDOGASTRAEA FAUST ЕТ TANG:
- Acanthocheilonema streptocerca (Macfie et Corson, 1922) Faust, 1949
- ГОД DIROFILARIA Dirofllarla loulsianensls (Faust, Thomas et Jones, 1941)
- ГОД MANSONFI) А Mansonella ozzardi (Man son, 1897) Faust, 1929
- КЛАСС «В СЕБЕ» И КЛАСС «ДЛЯ СЕБЯ»
- 1.9.2. Замкнутые классы
- Классы глаголов
- Важнейшие замкнутые классы
- § 21. Согласовательные классы
- 6.45. Понятие классов глагола
- Брачные классы.
- § 118. Классы глаголов
- Средний класс
- Рабочий класс
- Класс
- § 2. Социальные классы
- Высший класс