Морфология и биология стадий развития
Экзоэритроцитарная шизогония. Спорозоиты, инокулируемые в кровяное русло позвоночного животного, обладают строением, типичным для соответствующих стадий жизненного цикла класса
споровиков (с.
90). Как уже отмечалось выше, они дают начало экзоэритроцнтарным шизонтам, строение и локализация которых в организме хозяина определяются особенностями конкретных видов. IСпорозоиты сохраняются в кровяном русле примерно в течение часа после заражения хозяина, однако при этом количество |их заметно уменьшается. Они проникают в клетки ретикулоэндотелия и печени. Внедрившись в клетку, спорозоит округляется и превращается в кріштозоита. В течение первых двух суток это, как правило, небольшое округлое или овальное тельце, размеры которого нс превышают 2—2,5 мкм. В дальнейшем наблюдается быстрый рост крпптозопта. Для многих форм характерно образование крупных вакуолей, которые, однако, у зрелых, готовых к размножению особен полностью исчезают. Размеры зрелых криптозоитов моіуг достигать нескольких десятков микрон, а форма их сильно варки- руст в зависимости от типа клеток, в которых они паразитируют. Криптозонты, развивающиеся в паренхиматозных клетках печени, чаще всего имеют неправильную округлую пли овальную форму; виды, поселяющиеся в эндотелии капилляров, обладают вытянутыми, колбасовидными шизонтами (рис. 55, Б). |
В одних случаях (например, у РІ. ѵіѵах) зрелый, ставший много- ядерным шнзонт дает начало множеству небольших одноядерных телец — мерозонтов. В других же шизонт вначале делится на нисколько довольно крупных частей, от которых уже в свою очередь отпочковываются мерозоиты. Именно таким способом размножаются крнптозоиты PI. falciparum, паразитирующего у человека (рис. 55, А). I
Количество циклов бесполого размножения в тканевых клетках варьирует у разных видов. Формы, паразитирующие у птиц, обладают двумя поколениями первичных экзоэрнтроцитарных шпзЬн- тов, п лишь вторая генерация паразитов даст начало мерозонтам, способным инвазировать эритроциты.
У видов, вызывающих малярию млекопитающих, в том числе и человека, имеется только один цикл первичной шизогонии, после чего мерозоиты в большинстве случаев поступают в кровь. Из криптозоитов плазмодиев млекопитающих образуется до нескольких десятков тысяч мерозои’ігов (РІ. ѵіѵах — около 10 000, PI. falciparum — 30 000 и т. п.). |Сроки развития первичных экзоэрнтроцитарных шизонтов, соответствующие скрытому латентному периоду заболевания, различаются у разных видов, по для каждого конкретного вида относительно постоянны. Появление первых мерозонтов в кровяном русле у РІ. ѵіѵах наблюдается через 8 дней после заражения, у PI. falciparum через 5,5 дней: Первичная шизогония плазмодиев птиц протекает несколько быстрее: так, развитие двух поколений шизоидов PL gallinaceum занимает всего 120 ч. |
Как уже отмечалось выше, большинство видов р. Plasmodium обладает вторичной экзоэритроцитарной шизогонией. I
Фанерозонты могут существовать длительное время (иногда годами) и служат постоянным источником новых мерозонтов, посту- дающих в кровь хозяина. Именно этим явлением обусловливается способность ряда видов плазмодиев (у человека, например, РІ. ѵіѵах) вызывать так называемые отдаленные рецидивы (с. 123). Исключение составляют лишь виды, объединяемые в иодр. Ьаѵв- гапіа: паразит человека PI. falciparum и паразит высших обезьян
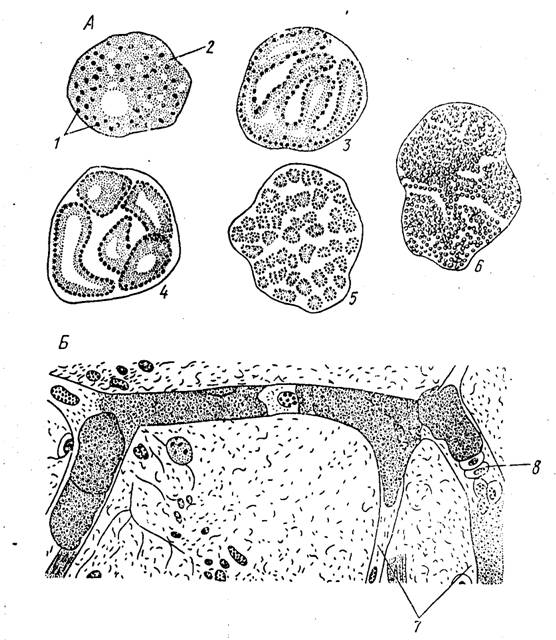
Рис. 55. Экзоэритроцитарпые шнзонты. А — последовательные стадии развития экзоэритроцитарного шизонта Plasmodium falciparum (по Гарнему); Б — вторичный экзоэритроцитарный шизонт Plasmodium gallinaceum в капиллярах головного мозга (по Брумпту):
/ — ядра шизонта, 2 — цитоплазма, 3—5 — последоаателыіыс стадии фрагментации шизонта, 6 — образование мероэонтов, 7 —капилляры головного мозг», 6—■ эритроциты
Рис. 56. Эндоэритроцитарная шизогония (из кн. Е. Нобля и Г.
Нобля). А—начало проникновения мерозоита в эритроцит; Б, В— последовательные стадии ’проникновения мерозоита; Г — молодой трофозоит в паразитофорион вакуоли; Д, Е—развитие трофозопта; Ж — начало шизогонии; 3 — скопление мерозонтов в эритроците; И — гамонт:/ — эритроцит, 2 — пелликула, 3 — микротрубочки, / — полярное кольцо. 5 —роптрии, 6 — ядро, 7 — двойные мембранные структуры, 8 — зернистыя эндорстнкулюм, У —гранулы гемозоипа, 10 — паразнтоформан иакуольPI. rcichenovi, у которых ІП'ОріІЧІКПІ зклоэрнтроцптарпая шизогония полностью отсутствует.
Своеобразной особенностью обладают виды плазмодиев, поражающие птиц. Фанерозоиты у них имеют двоякое происхождение: ;как и у видов из млекопитающих, часть из них представляет собой прямых потомков первичных шнзоитов, сохранивших способность развиваться в тканях, другая же часть возникает вторично из мсро- зоитон, поступающих из кровяного русла обратно в ткани внутренних органом. У плазмодием млекопитающих подобная картина никогда не наблюдается.
Эндоэритроцитарные шмзоптм. Большая часть мсрозоитов, образующихся из тканевых шизоптов, ппвазирует эритроциты и дает начало целому ряду сменяющих друг друга поколений эритроцитарных шизоптов (рис. 56). Процесс проникновения паразитов в клетки хозяина очень своеобразен. Мерозоит своим передним концом прикрепляется к эритроциту. Это соединение обеспечивается различием поверхностного заряда их мембран: по отношению к отрицательно заряженной мембране эритроцита пелликула меро- зонта ведет себя как обладающая положительным зарядом. После этого и начинается собственно внедрение паразита. Поверхностная мембрана красного кровяного тельца не разрушается, как это происходит при внедрении в клетки жппотпого-хозяина мерозонтов кокциднй (с. 88), а впячивается. Мерозоит погружается в образующееся углубление и в конце концов оказывается заключенным внутри замкнутой паразитофорпой вакуоли (рис. 56, А—Г). Мембрана последней таким образом берет свое начало непосредственно от поверхностной мембраны клетки хозяина.
В большинстве случаев в эритроците развивается лишь один трофозоит.
У ряда видов, однако, довольно часто наблюдаются случаи множественного заражения клеток хозяина, когда в эритроците поселяется сразу несколько (2—5) паразитов. Среди плазмодиев человека это явление чаще других наблюдается у PI. falciparum.Превращение мерозопта, имеющего довольно сложное строение, в трофозоит сопровождается существенными изменениями. Паразит округляется, размеры его не превышают 1—2 мкм. Почти сразу же около ядра появляется крупная вакуоль, которая быстро увеличивается в размерах. Ядро при этом смещается на периферию, а сама цитоплазма приобретает вид тонкого ободка (рис. 57, А). Эта очень характерная фаза развития трофозоита получила название стадии кольца. Дальнейший рост паразита сопровождается образованием многочисленных псевдоподий. Трофозоит становится очень подвижным (особенно у вида PI. vivax). Эта стадия часто обозначается как амебовидный шнзонт или амебоидный зародыш.
В цитоплазме паразита появляются первые гранулы малярийного пигмента — гемозоииа (с. 113), количество которых со временем неуклонно возрастает. Последующие изменения трофозоита сводятся к постепенному исчезновению вакуоли и втягиванию псевдоподий. Он увеличивается в размерах и занимает большую часть объема клетки хозяина. В этот период начинается деление
і
ядра. Число образующихся дочерних ядер различно у разных видов плазмодиев (12—18 — РІ. ѵіѵах\ 6—12, чаще 8,— PI. malari'ae\ 12—24, чаще около 16 — PI. falciparum и т. д.). Вокруг ядер обособляются участки цитоплазмы: это и есть процесс формирования нового поколения мерозоитов. Остается небольшое остаточное тело, содержащее многочисленные гранулы гемозоина. Мерозоиты е'ще некоторое время располагаются внутри клетки хозяина в виде компактного скопления (рис. 57, В). Разрушение эритроцита приводит к освобождению мерозоитов. Последние вместе с остатками
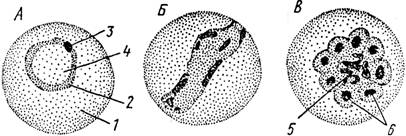
Рис. 57. Эмдоэріітроцнтармая шизогония Plasmodium mala- гіис (и і км. Дофлоіімп и Pcfixciioua).
Л—стадия «кольца»; її — стадия «амебоидного зародыша»; В — скопление мерозоитов в эритроците:/ — эритроцит, 2 — цитоплазма, 3 — ядро, 4 — пищеварительная вакуоль, 5—гранулы гемозоина, мерозонты
клеток хозяина и гранулами пигмента поступают в кровяное русло. Такова классическая картина развития трофозоита и процесса эндоэритроцитарной шизогонии, которую удается наблюдать на светооптическом уровне.
За последние годы, однако, накопилось много данных по ультратонкому строению трофозоитов и нх физиологии, позволивших выявить целый ряд особенностей кровяных споровиков, свойственных им как очень специализированным внутриклеточным паразитам.
По своему строению клетка трофозоита на первый взгляд мало отличается от клеток других простейших. Снаружи опа одета поверхностной мембраной, под которой непосредственно располагается цитоплазма. Трофозоиты обладают микропорой, столь характерной для всех споровиков (рис. 58, Л). В строении ядра плазмодиев, паразитирующих в птицах и млекопитающих, имеются существенные различия. У первых ядро всегда обладает хорошо выражіен- пой пуклеолой, тогда как вторые полностью лишены ее. В цитоплазме'клетки имеется зернистый эндоретикулюм и большое количество свободных рнбосомоподобных частиц. Характерные для простейших митохондрии с трубчатымгг кристами имеются только у плазмодиев птиц. Виды, паразитирующие в млекопитающих, обладают особыми мембранными образованиями, которым и приписывается роль митохондрий (рис. 58, Б). Во-первых, это так называемые двойные мембранные структуры, представленные двумя замкнутыми, вло-
женными одна в другую мембранами вытянутой, овальной пли округлой формы. Во-вторых, это концентрические полпмембраиные структуры. Последние несколько напоминают мсзосомы — примитивные мембранные образования бактерии, в которых содержатся, однако, типичные митохондриальные ферменты.
Питание трофозоитов осуществляется главным образом за счет цитоплазмы клетки хозяина.
Паразиты фагоцитируют цитоплазму эритроцита, в результате чего образуются крупные пищеварительные вакуоли. Самая первая пищеварительная вакуоль обычно является и самой большой по объему. Именно ее можно наблюдать под световым микроскопом у паразитов, находящихся па стадии «кольца». Заглатывание стромы эритроцита, пэ-вндпмому, может происходить как через микропору, так и в любой точке поверхности трофозоита.Основным источником энергии для паразитов является глюкоза пли близкие к пей сахара, поступающие из сыворотки крови жпвот- ного-хозяипа. Потребность в глюкозе эритроцитов, несущих тро- фозонты, в 25—100 раз превышает аналогичные потребности пепи- фицированных клеток. Большая часть поглощаемой глюкозы утилизируется, однако, самим эритроцитом, в котором накапливается ДТФ. Тем не менее, как показали специальные исследовании, паразиты способны использовать эту резервную ДТФ клетки хозяина для удрвлетворения собственных энергетических потребностей.
Глюкоза, используемая непосредственно самим паразитом, подвергается гликолитическому расщеплению. Значительная часть образующейся при этом пировиноградной кислоты превращается в молочную кислоту и лишь частично включается в цикл Кребса н окисляется до С02 и воды. Так как трофозонты до молочной кислоты расщепляют в среднем 4—6 молекул глюкозы и лишь одну окисляют полностью, то выход энергии за счет гликолиза и за счет цикла Кребса оказывается примерно одинаковым.
Однако, как показали специальные исследования, использование продуктов гликолиза в цикле Кребса может происходить только в том случае, если паразит находится внутри эритроцита. Это связано с неспособностью плазмодиев самостоятельно синтезировать особое вещество — кофермент А (КоА), без участия которого нормальный ход углеводного обмена оказывается невозможным. По-виднмому, трофозонты вынуждены использовать КоА, синтезируемый клетками хозяина. Уже только один этот биохимический «дефект» делает плазмодиев облигатными внутриклеточными паразитами.
Окисление глюкозы в' цикле Кребса теснейшим образом связано с дыханием паразита, которое осуществляется с участием цитохромной системы ферментов. Об этом говорит ингибирующее действие цианидов (специфических ингибитором цптохромов) па аэробное окисление энергетических веществ трофозоитами.
Потребность последних в аминокислотах, необходимых для синтеза белков, удовлетворяется за счет нескольких источников. Главным из них, по-видимому, является фагоцитируемый плазмодием гемоглобин, который, однако, утилизируется паразитом только частично.
Долгое время считалось, что малярийный плазмодий расщепляет молекулу-гемоглобина на две составные части: белковую — глобин и железосодержащую — гематин (железопорфирин), первый из которых гидролизуется до свободных аминокислот, второй же в виде коричпевато-бурых гранул накапливается в пищеварительных вакуолях. Однако, как показал специальный анализ, ьти пигментные гранулы не могут быть идентифицированы с гематином Малярийный пигмент, получивший название гемозонпа, обладаеч значительно большим молекулярным весом, содержит пептиды полностью отсутствующие в гематине, и в отличие от последнегс не ингибирует ферменты цикла Кребса (в частности, сукцнндегид- рогеназу). Гемозонп рассматривается как продукт частичной денатурации гемоглобина, вызываемой действием специальных протеолитических ферментов, продуцируемых плазмодием.
Из сыворотки крови трофозонты получают всего лишь одну аминокислоту—метионин. И, наконец, ряд аминокислот (аспарагиновая кислота, алапии и т. п.) плазмодии способны синтезировать сами.
О липидном обмене трофозоптов известно очень немного. Количество лшшдов и клетке паразита весьма значительно (до 30% сухой массы). По-видимому, наиболее активно паразиты синтезируют фосфолипиды.
Активное питание паразита, соответствующее периоду быстрой его роста, прекращается незадолго до начала шизогонии. Как уж< отмечалось выше, трофозонты в это время уже не образуют псевдо подии и перестают заглатывать гемоглобин. Шизогония начинаете! с быстрой фрагментации ядра. Этому процессу предшествует весь ма растянутый во времени период синтеза ДНК: он начинаете! еще на стадии кольца и заканчивается лишь у зрелого, готовом к делению паразита. Количество ДНК возрастает в 10—20 раз, чт< соответствует числу образующихся дочерних ядер, между которым: и происходит распределение гаплоидных наборов хромосом.
Процессы синтеза нуклеиновых кислот трофозоитамн обладаю некоторыми характерными особенностями. Эти специализированны внутриклеточные паразиты могут самостоятельно синтезироват только пиримидиновые основания (цитозин, тимин и урацил), тог да как их способность к синтезу пуриновых оснований (аденнн и гуанина) весьма ограничена. Большую их часть паразит в гото вом виде получает из клетки хозяина.
Ядра, образовавшиеся в результате деления, располагаютс; по периферии клетки. АДежду ними возникают глубокие впячпваіш: поверхностной мембраны, тик что формирующиеся мерозонты ока зываются почти полностью отдел сипы.ми друг от друга (см рис. 5U, Ж). Именно в этот период в них закладываются характер ные для этой стадии органоиды, отсутствовавшие у трофозоита Центральная безъядерная часть материнской клетки, в котороі скапливаются гранулы гемозоина, представляет собой остаточноїтело. После отделения мерозоитов от остаточного тела эритроцит, который к этому времени приобретает вид узкой каемки, окружающей паразитофорную вакуоль, окончательно разрушается.
Продолжительность развития трофозоитов между двумя периодами бесполого размножения различна у разных видов плазмодиев, по в пределах одного вида характеризуется высокой степенью постоянства. У большинства возбудителей малярии птиц этот срок составляет 24 ч, у PI. gallinaceum — 36 ч. Для видов, паразитирующих в высших обезьянах и человеке, как правило, требуется больше времени для завершения эритроцитарной шизогонии: РІ. ѵіѵах и PI. falciparum — 48 ч, PI. malariae — 72 ч и т. и. Характерной особенностью паразитирования плазмодиев в эритроцитах является удивительная синхронності» их развития, приводящая к почти одновременному формированию п выходу большого числа мсриаиптов в кровяное русло жниотпого-хозяппа.
Взаимоотношения трофозоита с клеткой хозяина сложны. Несомненно, однако, что влияние паразита на эритроцит очень велико. Примером может служить судьба мембраны, ограничивающей паразитофорную вакуоль и являющейся производным поверхностной мембраны эритроцита. Под действием трофозоита опа резко меняет свои свойства и превращается в образование, обеспечивающее транспорт некоторых веществ (например, ЛТФ), необходимых паразиту, из окружающей цитоплазмы. Характер локализации соответствующих ферментов говорит о том, что изменяется «полярность» мембраны, которая, по сути дела, функционально становится «частью» самого паразита.
С паразитофорной мембраной также, по-видимому, связаны особые образования, о существовании которых было известно очень давно. При окраске по Гимза-Романовскому в цитоплазме инфицированных эритроцитов выявляются небольшие участки, имеющие вид мелких точек, пятен и т. п. Для каждого вида плазмодиев форма, размеры, количество этих структур — довольно постоянные признаки. Это позволило использовать их в диагностических целях и присвоить им специальные названия: зернистость Шюффиера, зернистость Цимана, пятнистость Маурера и т. д. Электрошю-мпк- роскопические исследования показали, что вес эти пятна и зерна представляют собой вытянутые в длину или сферические участки, ограниченные параллельными или концентрическими мембранами. Во многих случаях они непосредственно связаны с мембранами, окружающими паразита (рис. 58, В). Высказывается предположение, что подобные структуры служат для увеличения площади поверхности, через которую осуществляется поступление пищевых веществ и выделение продуктов обмена.
Мёрозоиты. Размеры мерозоитов малярийных плазмодиев не превышают 1,5—2 мкм. Экзоэритроцитарные и эндоэрнтроцптар- ные мерозоиты несколько отличаются друг от друга по форме тела (рис. 59). Первые вытянуты в длину, тогда как вторые почти шаровидны. Снаружи клетку одевает пелликула, состоящая из поверхностной мембраны и двух подстилающих мембран. Под пелликулой, как и у мерозоитов кокцндий, располагаются микротрубочки. Передний конец несет так называемые полярные кольца, соответствующие прекопондалыіым кольцам кокцндий. В отличие от грега- рин н кокцндий мерозоиты плазмодиев полностью лишены коноида (с. 91). Имеются хорошо ныражсчшыс роптрии п мнкроисмы. Крупное ядро занимает центральную часть клетки. Цитоплазма
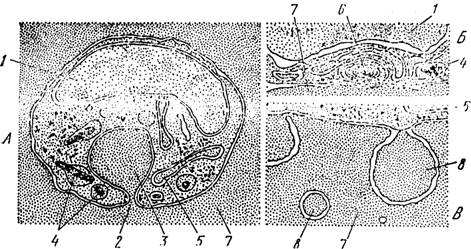
Рис. 58,, Тонкое строение трофозоита Plasmodium coalneyi, схема (по РудзнискоГі и Тригеру). А — молодой трофозонт; Л —.мембранные структуры трофозоита; В — мембранные структуры и цитоплазме эритроцита:
/ — ядро, 2 — микропора, 3 — пищеварительная вакуоль, 4 — гранулы гемо- зонна, 5 —двойные мембранные структуры, 6 — концентрические полимем* бранные структуры, 7 —цитоплазма эритроцита, 5—мембранные структуры \ в цитоплазме эритроцита
пронизана каналами зернистого эпдоретнкулюма. У мерозоитов плазмодиев, паразитирующих в птицах, в заднем конце располагается крупная митохондрия. В средней части клетки находится микропора. Превращение мерозоита в трофозонт после проникновения в эритроцит сопровождается исчезновением роптрпй, внутренних пелликулнрпых мембран, микротрубочек и полярных колец.
Дальнейшая еудьба мерозоитов различна. Чаеть пз них даеі начало новым поколениям трофозоитов, другая же часть превращается в гамонтов.
Половой процесс, спорогония .
Макро- и м и к р о г а м о н т ы. В большинстве случаев макрО' и микрогамонты у представителей р. Plasmodium появляются только после нескольких (минимум 1—2) циклов бесполого размножения. На ранних стадиях развития они весьма напоминают трофо- зонты, однако позднее различия становятся совершенно очевидными. Зрелые гамонты представляют собой в большинстве случаев округ-j лые или овальные тела, лишенные псевдоподий и вакуолей. Цитоплазма макрогамонтов окрашивается более интенсивно; чем у мик- рогамонтов. Имеются характерные различия и в строении ядер: у первых эксцентрично расположенное ядро компактно п обладает сравнительно небольшими размерами ('/в—Ѵю диаметра паразита), тогда как у вторых оно более рыхлое и крупное ('/j—7з диаметра клетки) и занимает центральное положение (рис. GO, Л, Ь).
Форма и размеры гамонтов довольно постоянны п пределах вида. Так, среди видов, паразитирующих в человеке, РІ. ѵіѵа.х

Рис. 59. Мсрозоиты малярийного плазмодия (по Порше-Хенне п Ви- пье). А — экзоэріітроціітарпый мс- розоит паразита птиц Plasmodium fallax\ Б — эндоэритроцнтарный меро- зонт:
/ — пелликула. 2—микротрубочки. 3 — ядро, 4 — митохондрия, 5 — зернистый эндо- ретикулюм, 6 — мнкропора, 7 — микроне- мы, 8 —роптрин, 9 —полярные кольца
Рис. СО. Га.чопты мгілирші- ного плазмодия. А — макро- глчоігг Plasmodium malarial”, Б — ммкроіамопт Plasmodium mulariad; ІЗ — .макро- гамонт Plasmodium falciparum-, Г--міікрипімоиг Plasmodium falciparum (Л, Б — по ДофлсГшу н Рейхеноьу;
В, Г— из кн. Грассе)и PI. ovale характеризуются крупными округлыми гамонтамн, занимающими весь объем эритроцита. Размеры последнего сильно увеличены по сравнению с незаражеииымн клетками. Гамопты РІ. та- Іапае имеют такую же форму, но значительно меньших размеров: присутствие паразита не вызывает заметного увеличения клетки хозяина. Очень своеобразной формы гамонты PI. falciparum, получившие название «полулунии Лаверана» (рис. GO, В, Г).
Гамонты р. Plasmodium обладают ограниченным сроком жизни, и поступление их в периферическую кровь жнвотного-хозяіша осуществляется за счет постоянно повторяющихся циклов бесполого размнеркения паразита.
В тонком строении гамонтов имеется много общего с другими стадиями жизненного цикла плазмодиев. В эритроците гамонты располагаются внутри иаразптофориой вакуоли. Так же, как н трофозоиты, они вызывают появление в цитоплазме клетки хозяина мембранных структур — зернистости Шюффеиа, пятнистости Маурера и т. п.
Гамонты одеты снаружи поверхностной мембраной, под которой располагаются внутренние мембраны пелликулы. Последние лучше развиты у мнкрогамонтов, тогда как у макрогамоптов они имеются лишь па отдельных небольших участках. Питание гамонтов на ранних стадиях осуществляется так же, как и у трофозоитов, путем образования пищеварительных вакуолей, в том числе и через хорошо развитую мнкронору. У зрелых гамонтов этот процесс, по- видимому, приостанавливается, так как пищеварительные вакуоли обнаружить не удается. Об интенсивности питания развивающихся паразитов может свидетельствовать большое количество гранул ге- мозоина в их цитоплазме.
Макро- и мнкрогамопты различаются по количеству рибосом и степени развития эндоплазматического рстпкулюма. У первых шероховатый ретикулюм хорошо развит и имеется большое-количество рибосом, тогда как у вторых они почти полностью отсутствуют. Полое интенсивная окраска цитоплазмы макрогамоптов па препаратах объясняется, по-видимому, именно этим обстоятельством. Кроме того, у макрогамоптов удалось еще обнаружить и мпк- ронемы.
Гамонты представляют собой последнюю фазу жизненного цикла малярийного плазмодия в организме позвоночного животного — все дальнейшее развитие паразитов осуществляется в переносчиках.
Гаметогенез и копуляция. Толчком к началу гаметогенези служит попадание зрелых гамонтов в пищеварительную' систему переносчика. Как уже отмечалось, переносчиками служат комары сем. Culicidae. Паразиты птиц чаще всего используют представителей р. Сиіех и Aedes. Переносчиками плазмодиев млекопитающих почти всегда являются комары р. Anopheles.
Молодые, еще не сформировавшиеся гамонты, а также прочие стадии развития паразитов, случайно попавшие в кишечник комара вместе с кровью, перевариваются хозяином. Зрелые же гамонты освобождаются из остатков эритроцитов и дают начало гаметам. Макрогамонты, по-видимому, не претерпевают никаких существенных изменений и целиком превращаются в макрогамету, сразу же готовую к оплодотворению.
Значительно сложнее протекает процесс микрогамстогснеза. Он начинается с того, что в цитоплазме мнкрогамонта в течение нескольких минут сначала образуются кннетосомы, а затем формируются и аксопсмы. Последние почти сразу же начинают производить активные ундулнрующие движения в толще цитоплазмы и передвигаются к поверхностной мембране клетки. Вслед за этим наблюдается деление ядра. 8 образовавшихся мелких ядер также мигрируют к поверхности клетки и располагаются рядом с аксоне- мами. Вокруг каждого такого комплекса обособляется участок клеточной мембраны. Возникает очень характерная фигура: крупное остаточное тело, от которого в разные стороны отходят 8 очень подвижных жгутовпдпых выростов (рис. 61). Полным отделением последних н завершается формирование микрогамет. Зрелая микрогамета имеет весьма простое строение. Это длинное жгутовидное 'Образование, в средней части которого сбоку от аксонемы располагается сильно вытянутое ядро.
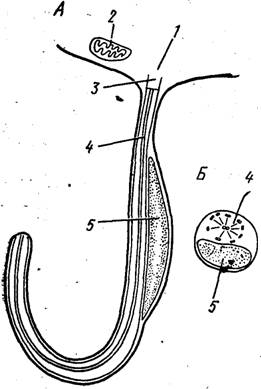
Рис. 61. Образование микрогаметы Plasmodium berghei (из кн. Гарнема). А — обособление зрелой микрогаметы от остаточного тела; Б — схема поперечного среза через микрогамету в области ядра;
1 — остаточное тело, 2 — митохондрия, 3 — кинетосома, 4 — аксонема, 5 — ядро
Подвижная микрогамета целиком проникает в макрогамету. Некоторое время в цитоплазме последней удается обнаружить как отдельно лежащее ядро микрогаметы, так и ее аксонему. В результате слияния ядер формируется диплоидная зигота, которая вскоре преобразуется в оокине- ту. Это особая подвижная стадия, способная к миграции в организме комара. Зрелая оокинета вытянута в длину и передний конец ее заострен (рис. 62,А). Имеется ряд органоидов, характерных для споровиков, — микротрубочки, микронемы и т. п.
Оокинеты внедряются в стенку кишечника комара. Иногда они проникают в клетки эпителия и располагаются в паразитофорных вакуолях. Однако довольно часто паразиты локализуются между эпителиальными клетками. В последнем случае развитие ооки- нет идет несколько быстрее.
верхность кишечника (рис. 62, £). заканчивается формированием спорозоитов.
О о ц и с т а, ф*о рмирование спорозоитов, спорозоиты. Молодая ооциста, одетая тонкой, но плотной оболочкой, по своей организации заметно отличается от оокинеты. Исчезают микротрубочки, микронемы. Начинается деление ядра, протекающее, по-видимому, очень своеобразно. Первый этап этого процесса •представляет собой эндомитоз, приводящий к значительному увеличению плоидности ядра. Последнее быстро увеличивается в размерах и становится лопастным, приобретая причудливую разветвленную форму. Ядерная оболочка, однако, сохраняет свою целостность. В дальнейшем ядро распадается на отдельные фрагменты, причем в каждом из них содержится гаплоидный набор генетическо-
В стенке кишечника переносчика происходит инцисти- рование оокинеты. На ее поверхности выделяется тонкая оболочка — циста. Паразит заметно увеличивается в объеме и постепенно начинает выпячиваться на наружную по- Это стадия ооцисты, которая
го материала. Наряду с мембранными структурами, столь характерными для трофозоитов плазмодиев из млекопитающих, появляются настоящие митохондрии. Огромное количество рибосом и изобилие РНК в цитоплазме ооцисты говорят об интенсивности процессов синтеза, протекающих на этой стадии развития паразита. Поступление пищевых веществ в ооцисту, по-видимому, осуществляет-
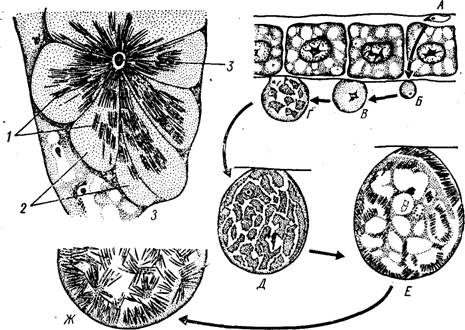
\у Рис. 62. Развитие ооцисты Plasmodium falciparum на стенке желудка комара (из кн. Грассе, с измен.). А — оокинета; Б — молодая ооци- ста; В—Д —^последовательные стадии развития ооцисты; Е — формирование спорозоитов в ооцисте; Ж — участок зрелой ооцисты со сформированными спорозоитами; 3 — спорозоиты в железистых клетках слюнной железы комара:
1 — спорозоиты, 2 — клетки слюнной железы, 3 — проток слюнной железы
ся через ее оболочку. Во всяком случае, последняя легко пропускает аминокислоты, в дальнейшем используемые паразитом для построения собственных белков. В ооцисте сохраняются также гранулы гемозоина и откладываются многочисленные капли жира. .
На поверхности цитоплазмы под наружной оболочкой появляются узкие и глубокие складки, что придает паразиту своеобразный «губчатый» вид.
Формирование спорозоитов начинается с того, что под поверхностной мембраной закладываются внутренние мембраны пелликулы, а немного позднее и микротрубочки (рис. 63). Они располагаются не сплошным слоем, а отдельными участками, которые вскоре начинают выпячиваться наружу. В образующуюся почку мигрируют ядро, митохондрии-и другие клеточные органоиды, формируются роптрии (рис. 63). В ооцисте развивается огромное коїшличество спорозоитов, которые располагаются вокруг одного или нескольких небольших остаточных тел с сильно вакуолнзирован- иой цитоплазмой.
Полностью сформированные спорозоиты, заполняющие полость зрелой ооцисты, по своему строению, очень сходны с мерозоитамп (рис. 64). Они обладают узким, сильно вытянутым в длину и слегка изогнутым телом, покрытым плотной пелликулой. Па самом переднем конце имеется небольшое углубление, по краям которого
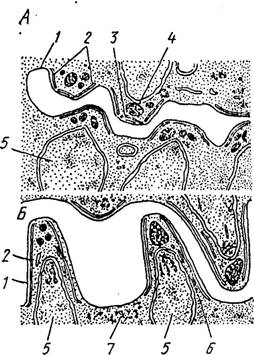
Рис. G3. Формирование спорозоитов в ооци- сте Plasmodium cynomolgi, схема (по Тер- закису). А, Б — последовательные стадии развития спорозоитов:
1 — поверхностная мембрана цитоплазматического тела ооцисты, 2 — закладка пелликулярных мембран. 3 — полярные кольца, 4 — роптріш, 5 — ядро, 6 — микротрубочки, 7 — остаточное тело
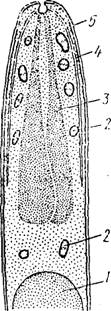
Рис. 04. Схема строения зрелого спорозоита Plasmodium galliiuiccuni (no Гарлему):
/ — ядро, 2 — мпкроиеми, 3 — роптріш, 4 — микротрубочки, 5 —пелликуларасположены полярные кольца. Коноид отсутствует. В передней половине клетки расположены хорошо развитые роптріш и микро- немы. Перед ядром, почти вплотную примыкая к нему, располагается аппарат Гольджи, а позади ядра — неправильной формы мп- тохЬндрин. В цитоплазме спорозоита имеются многочисленные рибосомы. Гистохимическл в ней выявляется большое количество РНК,- что можно рассматривать как своего рода преадаптацню к паразитированию в позвоночном хозяине: превращение спорозоита в экзоэритроцитарный шизонт сопровождается очень быстрым ростом, который просто невозможен без интенсивного синтеза белков.
Оболочка зрелой ооцисты, содержащей сформированные спорозонты и небольшое остаточное тело, сильно истончается. Такая
ооцнста легко разрывается, спорозоиты при этом попадают в гемо- лимфу комара. Они мигрируют по гемоцелю хозяина в передний конец тела и постепенно скапливаются в слюнных железах (см. рис. 62,-3). Большое количество паразитов проникает непосредственно в железистые клетки, в которых они располагаются радиально вокруг протока. Из клеток они попадают в проток, откуда уже при укусе комаром позвоночного животного инокулируются в кровяное русло последнего.
Малярия — опасное заболевание человека
У человека, как уже отмечалось выше, паразитируют 4 виде рода Plasmodium-. PI. vivax, PI. malariae, PI. falciparum и PI. ovate Заболевания, вызываемые ими, известны под общим .названием малярия, хотя между ними имеются некоторые существенные различия. PI. vivax — возбудитель так называемой трсхдисвной маля рни (malaria terliana), PI. malariae — возбудитель чстырехдиевноі малярии (malaria quartana); тропическая, или злокачественная малярия (malaria tropica) вызывается Pi. falciparum и, наконец PI. ovale вызывает малярию типа трехдиевной. Перечисленные выше виды отличаются друг от друга рядом морфологических признаков и биологических особенностей. Кроме того, каждый ИЗ ЭТИ) видов представлен большим количеством морфологически идеи тнчных штаммов, для которых характерны, однако, различна! степень патогенности, сроки развития в животпых-хозясвах, чувст иптелыюсть к химиотерапевтическим препаратам п т. п.
Между моментом инвазии человека спорозоитами п псрвымі клиническими проявлениями заболевания протекает известны! промежуток времени — инкубационный период. Длительность по следпего определяется, главным образом, сроками развития пер внчних экзоэритроцптарных шизонтов (8—12 дней у PI. vivax 5—6 дней у PI. falciparum и т. д.). Иногда, однако, в предела: одного вида встречаются штаммы, очень сильно отлнчающиес: друг от друга по продолжительности инкубации. Так, например у PI. vivax наряду с формами, развитие которых в печени человек; идет всего 8—12 дней (короткий тип инкубации), ^встречаются -штаммы, для которых характерен весьма длительный период нику бацни — до 6 месяцев. Первые приурочены к южным районам, зон; распространения вторых — области с умеренным климатом. ГІо-іш дпмому, такое различие в продолжительности инкубации може1 рассматриваться как адаптация к обитанию в разных кли,матиме скнх зонах — в северных районах, где лето начинается достаточт поздно іі сравнительно коротко, непродолжительный срок инкуба ции биологически невыгоден. Гамопты в кровяном русле хозяин, начнут появляться лишь в конце летнего сезона, когда количеств; комаров заметно снижается. Вероятность успешного завершенії: цикла при этом очень невелика. При длительном типе инкубаціи гамопты в крови человека появятся лишь в следующий летний се зон, по зато не в конце, а в первой половине, когда численності комаров достаточно велика, а высокие среднесуточные температуры позволяют паразиту успешно развиваться в переносчике.
После окончания инкубационного периода у больного начинает подниматься температура п появляются первые признаки лихорадки. Однако в начале заболевания колебания температуры еще не обладают строгой периодичностью. Это обусловлено неодновременным развитием первичных экзоэритроцптарпых шизоптов, в результате чего мерозоиты попадают в кровяное русло отдельными сравнительно небольшими порциями. На мазках крови, взятых у больного в это время, можно обнаружить паразитов, находящихся на самых разных стадиях развития. В результате первые генерации эритроцитарных мерозоитов выходят из разрушающихся эритроцитов неодновременно. Постепенное увеличение числа паразитов приводит к приобретению организмом иммунитета. Количество антител в крови больного быстро растет. Развивающийся иммунитет вызывает гибель отдельных небольших партий мерозоитов, что, в свою очередь, приводит ко все большей и большей синхронизации развития выживающих паразитов по мере повышения уровня иммунологического барьера. Так заболевание постепенно приобретает характер «правильной» лихорадки. Продолжительность промежутка, протекающего между двумя приступами, определяется временем, необходимым для развития трофозоптов п последующей шизогонии (с. 110). При заражении РІ. ѵіѵах и Pi. ovule приступы наступают через 48 ч, а при PI. malariae — через 72 ч. Наиболее остро протекает малярия, вызываемая PI. falciparum: приступы часто сильно растянуты и плохо отграничены один от другого. Состояние больного, страдающего тропической малярией, все время остается достаточно тяжелым.
Малярийный приступ начинается обычно сильным ознобом, который сменяется быстрым подъемом температуры. Приступы, особенно в случае тропической малярии, сопровождаются головными болями, общей слабостью, тошнотой, рвотами и т. и. 13 конце приступа наблюдается резкое снижение температуры и обильное выделение пота. Состояние больного улучшается. Характерная особенность малярии (кроме заражения PI. ovale)—строган приуроченность приступов к нерпой половине суток: наиболее тяжелое состояние больного приходится па утренние часы.
Приступ, сопровождающийся повышением температуры (рис. 65), является мощной ответной реакцией организма на поступление в кровяное русло в течение короткого промежутка времени большой массы мерозоитов, малярийного пигмента — гемо- зоппа, свободного гемоглобина, «обломков» эритроцитов и т. д. Для удаления всего этого инородного материала используются лимфоидные и ретикулярные клетки. Увеличение нечепи п селезенки, наблюдающееся при малярии, в первую очередь связано с усиленным размножением именно этих клеточных элементов и приростом ретнкуло-эндотелналыюй ткапн.
Весьма характерно для течения малярии и прогрессирующее малокровие, которое развивается, однако, нс только в результате массовой гибели пивазированных эритроцитов. Иногда, особенно при заражении человека тропической малярией, наблюдается явление гиперсепсибилизацни. Организм начинает вырабатывать аутоанті пол а. ГІоямлешіе последних приводит к агглютинации и гемолизу непораженных эритроцитов. Б плазму крови поступает огромное количество свободного гемоглобина, причем одним из последствий этого, кроме усиления малокровия, может быть час-
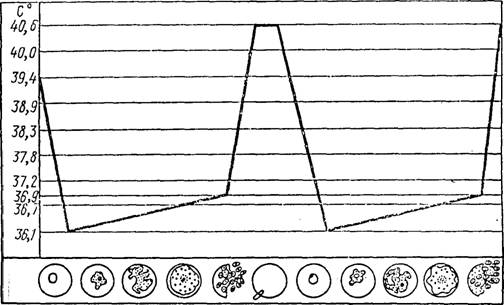
Рлс. 65. Температурная кривая больного, страдающего малярией, и се связц і
! с процессом размножения паразита в кровяном русле (нз кн. Грассе) !
І ;
тпчпая или полная блокада почек. В этом случае летальный исход иногда наступает' в течение 2—3 дней.
Не менее тяжелой формой протекания тропической малярии является развитие малярийной комы, сопровождающееся потерей сознания и нарушением рефлекторной деятельности организма Причиной комы, по-впднмому, являются закупорка капнллярот головного мозга инфицированными эритроцитами п нарушение проницаемости стенок сосудов. В этих участках иногда наблюдается дегенерация мозговой ткани. Кома, как правило, приводит к смерти больного.
Один нз характерных признаков малярии — ее рецидивирующее течение. По времени, протекающему между первичным прояв лопнем болезни її последующими рецидивами, различают два тип* последним ближние рецидивы, развитие которых' наблюдается і течение б месяцев после заражения, п отдаленные, наступающие і следующем сезоне, т. е. через год и более. Эти два типа рецнднвоі имеют различную биологическую основу.
С момента поступления паразитов в кровяное русло в крові больного появляются антитела и вырабатывается иммунитет. Сна чала это приводит, как уже отмечалось выше, к синхронизации ; развития эндоэритроцитарных стадий, а позднее и к почти полному і прекращению их размножения. Клинические картины, свойственные малярии, постепенно исчезают; наступает видимое выздоровление больного, в крови которого, однако, остается небольшое количество паразитов. Последние при ослаблении иммунологического барьера вновь начинают интенсивно размножаться, что сопровождается очередной вспышкой заболевания. Таким образом, через какой-то промежуток времени (от нескольких недель до нескольких месяцев) развивается ближний рецидив. Такого типа рецидивы, постепенно затухая, повторяются несколько раз. При доброкачественном течении малярии, не сопровождающимся какими-либо серьезными осложнениями, больной в конце концов полностью выздоравливает.
Отдаленные рецидивы имеют другую природу. Они обусловлены сохранением в клетках печени вторичных экзоэритроцптарных інпзонтон (с. 108). Под действием приобретенного иммунитета паразиты со временем синеем исчезают из кровяного русла, в то же время экзоэрнтроцптарпые стадии не подвергаются действию иммунных механизмов. С утратой или при значительном ослаблении иммунитета за счет этих шизоптов возобновляется поступление мероЗоитов в кровь, и болезнь вспыхивает с повой силой.
Ближние рецидивы характерны для всех форм малярии человека, отдаленные же могут иметь место только у тех видов, у которых сохраняется вторичная экзоэрптроцитарная шизогония. Именно этим обстоятельством объясняется'отсутствие отдаленных рецидивов при заражении тропической малярией (Plasmodium falciparum), тогда как для трехдневиоп малярии (РІ. ѵіѵах) они весьма типичны.
Распространение малярии. Малярия была и остается одним из самых распространенных заболеваний человека на земном шаре. Ареал малярии захватывает все климатические пояса, за исключением арктического. Наиболее широко встречающимся видом является возбудитель трехдневной малярии — РІ. ѵіѵах, самый ограниченный ареал у возбудителя PI. ovale, приуроченный к тропической Африке. Целенаправленная борьба с малярией привела за последние годы к значительному сужению границ ее распространения. Во многих районах, ранее считавшихся традиционно малярийными, это заболевание полностью ликвидировано. Наиболее показательны в этом отношении достижения Советского Союза. В начале века па территории России количество заболевавших малярией людей устойчиво держалось на уровне 3,5 млн. человек ежегодно. Несмотря .па две мировые войны и трудности послевоенного экономического развития, планомерное проведение противомалярийных мероприятий привело к значительному снижению заболеваемости населения: в 1949 г. число больных малярией сократилось до 1 0G3 53G человек. 19С0-Й год можно считать годом полной ликвидации малярии на всей территории Советского Союза. Сейчас ежегодное количество заболеваний не превышает нескольких де-
сятков, причем в основном это случаи, когда малярия бывает завезена из-за рубежа. I
Успешное наступление на малярию разворачивается в настоящее время но всему миру. Практически полностью завершено уничтожение малярийных очагов в странах Европы и Северной Америки. Малая Азия, Ближний-Восток, Индостан, Центральная .Америка — районы земного шара, в которых малярия еще встречается, однако благодаря широкой системе противомалярийных мероприятии ликвидация ее — дело ближайшего будущего. I
Однако и до сих пор имеются обширные территории, на которых борьба с малярией только разворачивается, где, по сути дела, сохраняются еще почти нетронутые очаги этого опасного заболевания. Юго-Восточная Азия, Центральная Африка, центральные районы Латинской Америки (в особенности огромная площадь, занимаемая бассейном р. Амазонки) и в наше время дают до 100 млн. случаен заболевания малярией в год. Причины такого положення имени нс столько биологический, сколько социальный характерні кроются н ткономпчсской отсталости государств, расположенных в перечисленных выше районах. |
Борьба с малярией. Борьба с малярией представляет собой сложный комплекс мероприятий, направленных на решение трёх основных задач: 1) воздействие на источник инвазии и борьба| с возбудителем, 2) борьба с переносчиком на всех фазах его развития и 3) защита населения от массовых нападений комаров. Только комплексная работа, охватывающая все эти три направления, могла привести п привела к победе над маля-рией. Воздействие на источник инвазии предусматривает, с одной стороны, тщательное п быстрое лечение уже заболевших людей, с другой же — массовые профилактические мероприятия для предупреждения распространения заболевания. ]
Набор препаратов, использовавшихся и используемых в настоящее время для лечения малярии, чрезвычайно широк. Одни из н'их подавляют процессы бесполого размножения паразитов в эритроцитах. Другие активно действуют на экзоэритроцитарные стадии, что особенно важно при лечении трехдневной малярии с отдалёнными рецидивами. Третьи вызывают гибель гамонтов. С эпидемиологической точки зрения их роль особенно велика. При уничтожении гамонтов в кровяном русле больного исчезает возможность заражения переносчиков, а следовательно, и'дальнейшего распространения малярнії. і
Наряду с активным лечением больных не менее важную роль играет массовая химиопрофилактика. В нашей стране, например, эти мероприятия широко проводились среди тех групп населения, которые подвергались реальной опасности заражения. |
Борьба с переносчиками малярии — комарами р. Anopheles также включает комплекс мероприятий. К ним относятся, с одной стороны, использование инсектицидов и биологических методов борьбы со взрослыми насекомыми и их личинками, с другой — широкое проведенію гидромелиоративных работ, обеспечивающих уіінчтоіке- шіе мест вынлода комаров. Радикальное оздоровление ряда районов в СССР (Закавказье, Средняя Азия, Белоруссия п т. д.) было достигнуто именно вторым путем.
Использованію различных химических веществ для борьбы с личинками комаров имеет давние традиции. Долгое время для этой цели применялись нефтевание водоемов, распыление различных ядов (парижская зелень, гексахлоран, ДДТ и т. п.). Весьма эффективными оказались н биологические методы борьбы — в частности, получившее широкое распространение заселение водоемов маленькой личинкоядиой рыбкой гамбузией. У нас в стране «гамбузирова- ние» водоемов успешно применялось в Закавказье и Средней Азии.
Внедрение в практику борьбы с насекомыми (и в частности с комарами) ДДТ ознаменовало крутой поворот в судьбе малярии как одного из самых пшрокораспространснных заболевании человека. Использование ДДТ позволило в очень короткий срок оздоровить огромные территории, на которых проживают сотнП миллионов людей. Оружие это оказалось, однако, обоюдоострым. Стойкость ДДТ и аналогичных препаратов приводит к тому, что при неумеренном использовании они длительное время сохраняются в природе и, что самое главное, накапливаются в живых организмах (в том числе и в организме человека), оказывая на них разрушительное действие. Наряду с этим комары приобретают устойчивость по отношению к инсектицидам,'что, естественно, резко снижает эффективность использования последних. Если в начальный период борьбы с малярией применение ДДТ и его аналогов было вполне оправдано, то в будущем главенствующая роль, несомненно, перейдет к биологическим и генетическим методам борьбы, не оказывающим столь катастрофического цоВдепствня па живую природу.
В уже существующих очагах заболевания наряду с мероприятиями, описанными выше, важное значение приобретает защита населения от нападения и укусов комаров. В свое время большую положительную роль сыграл метод зоопрофилактики. Суть его заключается в том, что между местами выплода комаров и жилыми помещениями располагается барьер в виде скотных дворов и хлевов. Домашние животные привлекают и задерживают значительную часть вылетающих па кормление самок. Важное значение имеют и методы индивидуальной защиты, из которых в последнее время наиболее широкое распространение получило использование репеллентов — веществ, отпугивающих комаров, и, в частности, диметилфталата.
Успехи в борьбе с малярией несомненны. Однако необходимо помнить, что малейшее ослабление внимания, преждевременное прекращение осуществления противомалярийных мероприятий в районах, где природные условия благоприятны для существования паразитов, могут привести к возникновению новых очагов заболевания.
Еще по теме Морфология и биология стадий развития:
- 2.6. Философские проблемы биологии и экологии[228] 2.6.1. Предмет философии биологии и его эволюция
- Понятие стадий развития в концепции Ж. Пиаже.
- Развитие познавательного интереса на уроках биологии
- 24. Развитие эволюционных идей в биологии и их значение для психологии.
- 1. Основные понятия морфологии как раздела грамматики. Морфология в системе грамматических дисциплин.
- Предмет и задачи морфологии. Связь морфологии с фонетикой, лексикой, словообразованием, синтаксисом.
- 33.Морфология. Лексич-ое и грам-ое значения в слове. Осн-ые понятия и ед-цы морфологии.
- 33.Морфология. Лексич-ое и грам-ое значения в слове. Осн-ые понятия и ед-цы морфологии.
- § 36. Предмет морфологии. Слово как объект морфологии
- Y.4.1. Концепция «Стадий экономического роста»
- 2.6.5. Воздействие биологии на формирование новых норм, установок и ориентаций культуры
- 2.6.2. Биология в контексте философии и методологии науки XX в.
- Значение стадий толкования
- 3.4. Модель последовательных стадий