. ПОДКЛАСС GREGARIN1NA DUFOUR — ГРЕГАРИНЫ
Грегарины — очень своеобразная группа споровиков, паразитирующих в различных беспозвоночных животных, главным образом в членистоногих. Большинство грегарин обитает в просвете кишечника своих хозяев.
Некоторые виды поселяются в полостях других внутренних органов и в полости тела. У многих грегарин обязателен период внутриклеточного паразитирования, который может быть, однако, очень кратковременным.Жизненный цикл подавляющего большинства представителен этого подкласса характеризуется отсутствием бесполого размножения— шизогонии и включает в себя только половой процесс и спорогонию. Лишь у небольшого числа видов шизогония имеет место. Развитие грегарин в этом случае осуществляется по обычной схеме, характерной для многих кокцнднй (с. 84). В строении грегарин теснейшим образом переплетаются признаки, свойственные всем споровикам, с признаками далеко зашедшей специализации.
Разработка естественной системы подкл. Gregarinina еще очень далека от своего завершения. Долгое время принималось, а иеко-
Рис. 6’8. Схема жизненного цикла Leucocytozoon siniondi (из кн. Грассе):
/ — микро- и макрогамонт в эритроцитах, 2 —■ образование микрогамет и макрогаметы, 3 — копуляции. 4—6 — миграция оокнисты, / — последовательные стадии развития ооцнеты, 4 — выход спорозоитов из ооцисты, 9 — внедрение мерозоита в эндотелиальную клетку, 10—12 — последовательные стадии развития экзоэрнтроцитарного шнзонта, 13 — выход мерозоитов в просвет кровеносного сосуда, 14 — эритроциты с проникшими в них мерозонтамиторыми исследователями принимается и до сих пор, деление подкласса на 2 отряда: Schizogregarinida, объединяющий виды, в жиз* ценном цикле которых имеется шизогония, и Eugregarinida (настоящие грегарины), у которых шизогония отсутствует.
В 1953 г. французский зоолог П. Грассе (Р.
Grasse) предложил новую систему, согласно которой грегарины объединяются в 3 отряда: Archigregarinida, Eugregarinida и Neogregarinida. С некоторыми уточнениями, внесенными позднее, эта система и принимается в настоящее время большинством протозоологов.Отр. Archigregarinida — архигрегарииы включает в себя наиболее примитивных грегарин, которые почти на всем протяжении своего развития сохраняют морфологические особенности, свойственные спорозоитам остальных споровиков. У отдельных впдое встречается шизогония. Паразитируют главным образом в кишечнике иолихет.
Отр. Eugregarinida — настоящие грегарины. Самый большой отряд, объединяющий виды, развитие которых всегда осуществляется без шизогонии. Паразитируют в различных беспозвоночных животных, но в основном приурочены к членистоногим. В состаі отряда входят такие широко распространенные роды, как Gregari- па, Monocyst is и др.
Отр. Neogregarinida — неогрегарины представлен сравнительна небольшим числом видов, обладающих бесполым размножением — шизогонией. Последняя, по-впдимому, возникла в жизненном ЦИКЛІ этих паразитов вторично. Встречаются только в насекомых.
Жизненные циклы грегарин
♦ І
• Наиболее распространенным и типичным может считаться жиз ненный цикл, свойственный представителям отр. Eugregarinidi (рис. 69,70). Инвазионным началом служит ооциста, содержащаі сформированные спорозоиты. При заглатывании ооцнеты живот ным хозяином освободившиеся из нее спорозоиты внедряются : стенки кишечника и в большинстве случаев переходят к внутрикле точному паразитированию. Однако эта стадия, как правило, очен непродолжительна и паразиты снова возвращаются в просвет.км шечішка. Спорозоиты тех видов, которые обитают в полости тел или в других органах хозяина, совершают миграцию к месту оков чатслыюй локализации.
Спорозоиты превращаются в гамонтов (с. 36). Интенсивн питающиеся и растущие гамонты часто называются трофозонтамі Первой стадией своеобразного полового процесса грегарин можн считать образование сизигия: две грегарины прочно соединяютс друг с другом и в таком виде могут довольно длительное врем существовать в хозяине, продолжая двигаться, питаться и расті Дальнейшие преобразования сизигия заключаются в том, что об особи округляются и, не теряя собственной целостности II инднві дуальности, выделяют на поверхность общую оболочку.
Эта стади называется гаметоцистой. Внутри гамстоцисты начинается формі отдельные ооцисты выходят во внешнюю среду по семяпроводам хозяина (рис. 70).Схема жизненного цикла представителей отр. Archigregarinida в подавляющем большинстве случаев не отличается от вышеприведенной. Лишь у некоторых, по-видимому, наиболее примитивных видов имеет место шизогония. В этом случае спорозоиты внедряются в толщу стенок кишечника и приступают к бесполому размножению. В результате образуются большие скопления мерозоитов.

Рис. 71. Жизненный цикл грегарин р. Selenidlutn (по Шревелю, с измен.І!
] спорозоит, 2 —гамоит, 3— сішігнП, 4 —начало образования гамотоцнсты, 5 — деление ядер гамонтов в гамстоцисте; 6 — образование гамет, 7 — мужская гамета* 8 — женская гамета, 2 —зигота, ІО—12 — последовательные стадии спорогонии
Последние и дают начало гамонтам, объединение которых в сизигий знаменует собой начало полового процесса (рис. 71).
Совсем иная картина наблюдается у представителей отр. Neo- gregarinida. Неогрегарины, будучи филогенетически связаны в своем происхождени с утратившими шизогонию настоящими гре- гарииами (с. 131), по-видимому, вторично приобрели бесполое раз» [4] [5]

множсние, несколько циклов которого у них обязательно предшествуют половому процессу (рис. 72). Спорозоиты этих грегарип таким образом превращаются не в гамоптов, а в пшзоптов. Судьба же мерозоитов может быть различной. Одни из них снопа дают начало бесполому поколению, другие превращаются в гамоптов и приступают к половому процессу.
Морфология и биология стадий жизненного цикла. Гамонты
Спорозоиты, освободившиеся из ооцнст в просвете кишечника животиого-хозяина, обладают строением, характерным для этой стадии жизненного цикла всех споровиков (с. 90). Превращение спорозоитов в гамонты в большинстве случаев сопровождается серьезной перестройкой их организации: утратой одних органоидов и развитием совершенно новых.
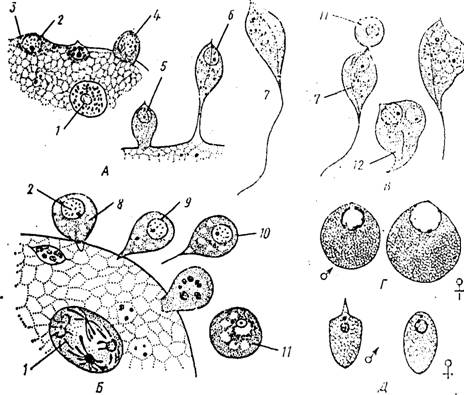
Многие виды грегарин в своем развитии проходят этап внутриклеточного паразитирования (рис.
73, Л). Спорозоиты внедряются в клетки эпителия кишечника, округляются п начинают расти. Постепенно увеличиваясь в размерах, они занимают значительную часть объема пораженной клетки. При последующем развитии большая часть тела формирующегося гамонта выходит из клетки хозяина и оказывается лежащей непосредственно в просвете кишечника. Лишь самый передний участок тела паразита остается закрепленным в стенке этого органа. Гамопт продолжает увеличиваться в размерах и приобретает черты строения, характерные для данного вида грегарин.Период внутриклеточного развития иногда может сильно удлиняться. Существуют виды (Lankesteria и др.), гамонты которых вплоть до начала полового процесса обитают в клетках хозяина (рис. 73, Б); по выходе из последних они сразу же приступают к образованию ензигиев. Часто, однако, наблюдается и противоположная картина. У некоторых грегарин спорозоиты не внедряются целиком в клетки эпителия кишечника, а только закрепляются в них передним концом (рис. 73, В). Все развитие паразита протекает, таким образом, виеклсточпо.
Строение зрелых гамоптов грегарип очень разнообразно. Наиболее примитивные виды, относящиеся к отр. Archigregan'nida, па протяжении всей своей активной жизни в просвете кишечника полихет сохраняют черты организации, свойственной спорозоитам, но достигают при этом очень больших размеров (до 300—400 мкм). Зрелые гамонты архигрегарин обладают вытянутым в длину телом, самый передний конец которого отделен от остальной части плав- ным-суженнем (рис. 74, А). Этот участок, с помощью которого паразиты могут закрепляться в эпителии кишечника, называется мукроном (mucron). Вдоль всего тела паразита тянутся широко расставленные неглубокие борозды (рис. 74, Б). Под хорошо развитой пелликулой, образованной, как и у других споровиков, тремя мембранами, располагаются многочисленные микротрубочки. Последние, по-видимому, обеспечивают подвижность паразіыов: ра-
Рис. 73. Развитие гамонтов.
А — развитие гамонта Gregarina fernandoi в эпителии кишечника таракана Pycnoscelus surinamensis (по Мано- Сабаратиам); Б — развитие гамонта Lankesteria clarki в эпителии кишечника'комара Aedes sierrensis (по Сандерсу и Пойнару); В — развитие гамонтов Gregarina garnhami в кишечнике саранчи Schisiocerca gregaria (по Харри):1—спорозоит, 2 — эшіліерпт, 3 — деЛтомсрпт, Ч — нротомернт, 5 — мукроп, £ — ядро эпителиально!! клетки, 7 — развивающие»! гамонтыстущие и зрелые гамонты способны активно изгибаться из стороны в сторону.
Передний конец —«мукрон (у развивающихся паразитов он обычно погружен в клетки эпителия кишечника хозяина) несет отчетливо выраженный коноид и 2—3 прекоиоидалышх кольца, кото-
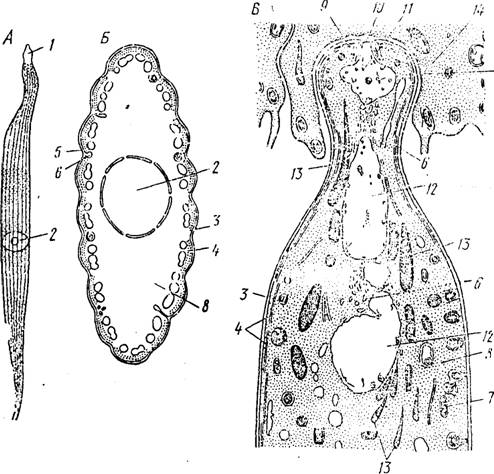
Рис. 74. Строение гамопта р. Selenidium. А — общий шід гамоита Sclenidium faushaldi (по Левайну); Б — схема поперечного разреза через тело гамон- та (по Вивье и Шревелю); В — схема строения переднего конца тела га-
монта (по Шревелю):
/ — мукрон, 2 «— ядро, J —пелликула, 4— микротрубочки, 5—микромора, £ — пппоцм- тозная вакуоль, 7 — митохондрия, 8 — эндоплазма, 9 — коноид, 10 — прекоиондаль- ные кольца, U — мукрональная вакуоль, 12 — пищеварительные вакуоли, ІЗ —рои- трни, 14 — клетка эпителия кишечника
рые, по-видимому, соответствуют аналогичным образованиям других споровиков (с. 90) (рис. 74, В). Сюда же направляются крупные многочисленные роптрии.
У примитивных грегарин коноид играет роль цнтостома, через который осуществляется поглощение пищевых веществ. Внутри коноида проходит широкий канал, на конце которого образуется крупная пищеварительная вакуоль. Обычно еще 2—3 такие же вакуоли располагаются в передней половине тела паразита. На их поверхности в свою очередь образуются многочисленные пиноци- тозные пузырьки, которые и поступают в эндоплазму паразита, Роптрии, по-видимому, участвуют как в лизисе эпителиальных клеток кишечника хозяина, так и в процессах внутриклеточного пищеварения.
В пользу последнего предположения говорит наличие в роптриях большого количества кислой фосфатазы и их тесный контакт с пищеварительными вакуолями. Важную роль в питании
Рис. 75. Различные типы гамонтов отр. Eugregarinida (из кн. Грассе). А — Monocystis proteiformis из олигохет; Б — Schneideria mucronata из личинок двукрылых); В — Menospara polyacantha из личинок стрекоз; Г — Taeniocystis тіга
Archigregarinicla,- вероятно, играют и микропоры, количество коте рых может быть весьма значительным. Они также принимают учг стие в образовании пнпоцнтозных вакуолей (рис. 7-1, Б).
В центральной части клетки располагается крупное пузырьке видное ядро. В цитоплазме разбросаны многочисленные мптохощ рии. Весьма значительно и количество аппаратов Гольджи. Хор< шо развит зернистый ретикулюм. Гранулы запасных питательны веществ — нарагликогспа заполняют практически всю эндоплазм паразита. По своей химической природе параглнкоген представлж собой полисахарид амилопектин, свойственный, по-видимому, другим споровикам.
Наибольшего разнообразия строение гамонтои достигает в отр. Eugregarinida (рис. 75). Самые примитивные представители этого отряда сохраняют определенные черты сходства с архнірегарппа- ми. Они обладают цельным нерасчлепениым телом, па переднем конце которого располагается отчетливо выраженный мукрон. Тонкое строение этого образования, однако, подвергается заметным изменениям. Коноид, а в некоторых случаях и роптрпп исчезают (рис. 76). Сохраняются лишь преконоидальиые кольца и огромная вакуоль, получившая название мукропалыюй. Опа, по-ипдпмому,

Рис. 76. Строение мукрона у представителей отр. Eugregarinida, схема.
А — передний конец тела гамонта Lankesteria barretti (но Вппра); ІЇ — прикрепление гамонта Lecudina pellucida к клетке эпителия кишечника при помощи мукрона (по Шревелю и Внвьс):
/ — пелликула, 2 — пелликуляриые гребни. 3 — поверхность мукрона, •/ — мукромлль- ная вакуоль, заполненная фибриллами, 5—ядро, С — гранулы іирлс.інкіиеіиі. 7 — роптрии, в —клетки эпителия кишечника хозяина, У —ядро эпителиальной клетки,
10 — участок измененной цитоплазмы эпителиальной клетки
соответствует пищеварительной вакуоли архнгрегарпп, хотя и отличается от последней целым рядом признаков. Участок поверхности тела, ограниченный преконоидалыіым кольцом, как правило, одет не пелликулой, а обычной поверхностной мембраной. Именно к нему н примыкает мукрональная вакуоль. Последняя при помощи системы каналов и перемычек соединяется еще с несколькими крупными вакуолями, расположенными глубже в эндоплазме паразита. От поверхностной мембраны мукрона в глубь этого сложного вакуолярного аппарата отходят тончайшие фибриллы, напоминающие по своей структуре мнонемы некоторых инфузорий.
У,видов, развитие гамоптов которых целиком протекает внутри клеток хозяина, мукрон так же, как и у Archigregannida, выполняет, по-видимому, трофическую функцию и служит для сорбции пищевых веществ. Иная картина наблюдается у полостных паразитов, например у представителей р. Lecudina, которые лишь удерживаются на эпителиальных клетках кишечника при помощи пе- редмсго конца тела. Их мукрон становится органоидом прикрепле имя. Его мягкая и эластичная поверхность используется паразнтої в качестве присоски. Сохраняется, но-шідпмому, п трофическа: функция, о чем говорит сильное изменение участка клетки, к кото рому прилегает мукрон. Цитоплазма клетки хозяина в этом мест вакуолнзнрустся, типичные клеточные органоиды исчезают, а в об ласти непосредственного контакта поверхностной мембраны клеткі с мукроном появляются многочисленные мелкие пузырьки (рис. 76 В). Все эти изменения могут быть вызваны.только действием сне цифпчеекпх энзимов, выделяемых паразитом..

Рис. 77. Различные типы эпимеритог) У настоящих грсгаріш (из кн. 1 Грассе). /1— S/ylocephatus longicollis', Б— Pyxinoidcs balani\ В — Corycella аГіпаіа:
/ — імшмерит, 2 — иротомерит, .‘і — дсПгоМеРі,т. 4 — ядро, 5 — клочки :лііполия
кшлечшіка
Подавляющее большинство настоящих грегарпп обладает н сколько иным строением. Они полностью утратили мукроп, а те. их отчетливо подразделяется на 2 или 3 отдела. Передний кон> грегарины преобразуется в так называемый эпимернт, при помои которого паразит постоянно или временно, только на определе ном этапе развития, закрепляется на стенке кишечника хозяин В отличие от мукрона эпимерит всегда четко отделен от остально тела грегарины глубокой перетяжкой и, по-видимому, не содерж никаких сократительных элементов- Прикрепление паразита К'Сте ке кишечника осуществляется только по принципу заякорнваш Форма эішмерига очень разнообразна. В одних случаях он мож быть шаровидным, в других вытягивается в длинный хоботок, п боко погружающийся в ткани хозяина, иногда эпимерит превраіг ется в настоящий «якорь», снабженный острыми направленны; назад зубцами (рис. 77). У молодых растущих грегаріш эпнмері по-видимому, обладает высокой метаболической активностью. ( этом говорит наличие в его цитоплазме большого количества риб
сом, аппаратов Гольджи, сложной вакуолярной системы и т. д. В литературе высказывается предположение о том, что па этой стадии развития эпимерит играет важную ролі, в поглощении пищевых веществ паразитом. Механизм этого процесса, однако, еще совершенно неясен.
Судьба эпимерита у разных видов грегарин различна. Иногда он сохраняется длительное время. Однако у очень многих видов

Рис. 78. Строение пеллнкулярных гребней гамонтов настоящих грегарин. А — схема строения поверхностного участка тела Lecudtna pellucida-, Б — схема движения пеллнкулярных гребней Lecudtna pellucida (по Впвье, с измен.); Б — схема работы пеллнкулярных гребней Pileuccphalus blaberae
(но Вайра):
/ — пелликула, 2 *— «базальная мембрана», 3 — эктоплазма, •/ —кольцевые фибриллы, 5 — митохондрия, 6 — гранулы парагликогена, 7 — капли жира, 6 — мпкропора,
9 — эндоплазма
переход к существованию в просвете кишечника сопровождается полной утратой этого образования (см. рис. 73, А). Следующее за эпимеритом тело гамонта может быть либо цельным (Schneide- ria, Stylocystis и др.; см. рис. 75, Б), либо состоит из двух отделов—протомерита и дейтомерита, что свойственно очень многим видам (см.-рис. 73, А, В; 75, В). Протомернт значительно меньше дейтомерита и отделен от последнего отчетливо видимой перегородкой—септой. У некоторых видов грегарин дентомерит может за счет образования неглубоких перетяжек разбиваться на несколько «члеников» (см. рнс. 75, Г).
Очень далеко заходящей специализации у всех представителей отр. Eiiprregarinida подвергаются поверхностные структуры клетки— пелликула и прилегающий к ней слой эктоплазмы. У боль-
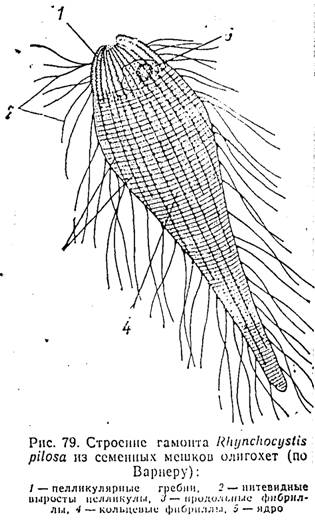
шжіства настоящих грегарин пелликула образует узкие и высокие гребни, тянущиеся ндолі. всего тела (рис. 78). В вершинной часті каждого такого гребня проходит по несколько продольных фиб рилл. Очень часто зона гребней отделяется от остальной эктоплаз мы хорошо выраженным плотным слоем — «базальной мембраной» Между гребнями располагаются продольные ряды многочисленны: микропор. Сублеллику- лярные микротрубочки у еугрегаринид, как правило, отсутствуют. Они заменяются системой расположенных в эктоплазме фибрилл, которым некоторые авторы приписывают сократительную функцию. Соответственно в литературе их часто называют мионемами. У ряда видов, паразитирующих в полости семенных мешков олигохет, эта фибриллярная система может достигать большой сложности п состоять как на продольных, так н кольцевых волокон (рис. 79). У многих кишечных паразитов чаще всего сохраняются только кольцевые фибриллы, залегающие в толще эктоплазмы (рис. 78,А).
Описанные выше особенности строения поверхностных образований гре- гарин из отр. ІІиугеуагі- nida позволяют попять механизмы питания и движения этих крайне специализированных простейших.
; Усвоение пищевых веществ молодыми гамонтами, как уже с мсчалось выше, ио-вмдимому, частично осуществляется за сч физиологически активного эпимерита. После отделения эпимери и перехода грегарины к жизни в полости кишечника этот пу поступления пищи в организм паразита становится невозможны Вероятно, как и у других споровиков, многочисленные МИКрОПО] представляют собой настоящие ультрацптостомы, служащие д поглощения ІШЩИ.
Много споров в научной литературе было связано с попытка: объяснить механизм очень своеобразного скользящего епосо движения, свойственного большому числу видов настоящих греі ріш, обитающих в кишечнике хозяина. Вез каких-либо видимых изменении формы тела (изгибании, скручивании и т. и.) простевшие медленно перемещаются в окружающей их среде.
Еще в конце прошлого века русский зоолог В. Т. Шевиков связывал этот тип движения грегарнн с выделением слизи через многочисленные норы в пелликуле. Стекая по узким бороздкам пелликулы к заднему концу тела, елнзв застывает здесь в виде топкого стебелька. Постоянно нарастая за счет все новых и новых порций слизи, стебелек проталкивает грегарппу вперед. Б. Ф. Соколов уже в начале XX в. модифицировал гипотезу Шевякова. Он считал, что слизь не просто вытекает из пор, а с силон выбрасывается от туда. При этом возникает реактивная тяга, обусловливающая перемещение организма.
Специальные наблюдения, проведенные позднее, показали, однако, что далеко по всегда скольжение сопровождается выделением слизи гамоитамп. Более того, искусственное разрушение слизи нагреванием пли удаление ее путем тщательного отмывания простейших почти не влияют ни на характер, ни на скорость движения. D этой связи было высказано предположение, что скольжение грегарнн возможно только при соприкосновении их с твердим субстратом. Оно является результатом перистальтического сокращения, обращенного к субстрату .участка поверхности тела, которое вызывается действием эктоплазматических миопем. Выделяемая же паразитом слизь лишь способствует уменьшению трения. Таким образом, в соответствии с этими взглядами механизм скользящего движения настоящих грегарнн оказался в какой-то мере сходным с механизмом движения брюхоногих моллюсков.
Широкое использование просветной и сканирующей электронной микроскопии позволило объяснить тонкий механизм для скользящего движения. Целый ряд исследователей (Ѵіѵіст, Ѵаѵга и др.) считают, что главная роль принадлежит упоминавшимся выше пелликулярным гребням. В одних случаях, как, например, у паразита полихет Lecudina pellucida, на поверхности зела грегарппы чередуются неподвижные гребни и гребни, способные к упдулирую- щему движению (рис. 78, Б). Благодаря действию последних в узких промежутках между неподвижными гребнями могут возникать токи жидкости, направленные к заднему концу тела, что и обеспечивает перемещение грегарины вперед. У большинства же видов грегарнн к ундуляции способны, по-видимому, все гребни. Движение двух соседних гребней всегда осуществляется в противоположных фазах (рис. 78, В), так что между ними возникают более или менее изолированные камеры. Последние, перемещаясь от переднего конца тела к заднему, проталкивают и содержащуюся в них жидкость, которой может быть как окружающая грегарппу среда (содержимое кишечника хозяина, культуральная жидкость и т. и.), так и слизь, продуцируемая самим паразитом. Для выделения слизи на поверхность тела, по-видимому, используется часть мпкропор. Описанный выше механизм хорошо объясняет способность грегарнн передвигаться в толще жидкости, а не только по твердому субстрату. Становится понятным и скользящее движение тех видов, которые выделяют мало слизи (например, упоминавшаяся выше L. pellucida). Ундуляция гребней, по-видимому, возникает в результате сокращения фибрилл, расположенных в вершинах гребней.
У видов, не обладающих способностью к скользящему движению, изменяется и строение поверхностных образований клетки. Примеров подобного рода особенно много среди представителен упоминавшегося выше ссм. Monocystidae — паразитов семенных мешков олпгохет. Иногда гребни у них сохраняются лини, па самом переднем конце тела, служащим для прикрепления паразитов к скоплениям развивающихся половых клеток; остальная же поверхность несет многочисленные тонкие волосовидные выросты (рис. 79).
Граница между экто- и эндоплазмой определяется зоной распространения многочисленных и разнообразных вакуолей и включений. Среди последних доминируют параглнкогсповыс гранулы количество которых может быть очень велико. У зрелых гамонтов они целиком заполняют всю эндоплазму, из-за чего под световым микроскопом эти простейшие кажутся совершенно непрозрачными, В эндоплазме же располагаются и многочисленные клеточные органоиды: аппараты Гольджи, каналы эндоретикулюма,. митохондрии. Крупное пузырьковидное ядро у форм, тело которых подразделяется на прото- и дсіітомерпт, всегда локализуется в последнем.
Гамонты небольшого специализированного отр. Neogregarinida имеют округлое или вытянутое тело, не расчлененное на отдельные участки. Они долгое время, вплоть до образования сизигия, сохраняют черты организации типичных споровиков. Передний конец гамонтов несет коноид, роптрни, мукрональную вакуоль, преконои- дальные кольца и т. д.
Половой процесс
Сизигии. Объединение гамонтов в сизигий представляет собой первый этап'полового процесса этих простейших. Форма спзигііев очень разнообразна. В большинстве случаев две грегарипы соединяются цепочкой: одна особь своим передним концом—эпнмери- том •—прикрепляется к заднему концу дейтомсрпта другой (рис. 80, А). Передняя грегарнна при этом обозначается как при-

Рас. 80. Сазаній грсгараа. А — Оге- garina sp. (аз ка. Грассц с измен.); Б — Stiiloccplialiis longicallis (аз кн. Грелля); В — i>'clenUlitun sabellariae (по LUjH'iie.i.iio):
J — іфимит, 2 — стеллит
мит, а задняя — как сателлит. Очень часто особи, образующие сизигий, соединяются передними концами, как это имеет место у многих Eugrcgarini- cla и Neogregarinida (рис. 80, Б). Для Archigregarinida, напротив, часто свойственно объединение грегарнн задними концами (см. рис. 71). И, наконец, у ряда видов при образовании сизигия особи прикрепляются друг к другу боковыми поверхностями тела (рис. 80,
В).
Сроки образования еизпгп- ев варьируют у разных видов. 13 тех случаях, когда все раз- витистамоіітов- протекает, вііутриклеточио, как, например, у представителей р. Lankesteria, сизигии формируются очень поздно, лишь после освобождения паразитов из клеток хозяина. Почти, сразу же происходит образование гамстоцисты. Иная картина, на?, блюдаётся у видов, которые рано переходят к существованью'в' просвете кишечника. И этом случае объединиться в сизпгіпі могут очень молодые незрелые гамопты, рост п развитие которых после этого еще продолжается длительное время.

' Рис. 81. Различные типы соединения гамоитои и сп.'шпш. .1 — Grega- rina polymorpha (по Дсвошсліо); В — Lintaroujstis fungoiilvs (но Депорту); В — Diplauxis hatli (но Вітье с сопит.):
I — задний конец тела принята, 2—-пелликула, 3 — неллпкулярпые гребни,
4 — разросшийся участок леллнкулярных гребней сателлита, .5 — «базальная мембрана», 6 — протомернт, 7 — дейтомерпт, Д — гранула парнілмкогаіа. 9 — аппарат Гольджи, 10 — ядро, // — каналы эпдоротикулюма
Прочное прикрепление одной особи к другой сопровождается серьезными изменениями поверхностных структур грегарин в зоне контакта. При образовании сизигия классического типа наибольшие перестройки затрагивают сателлит (рис. 81, Л). Па переднем конце его протомерита происходит сильное разрастание зоны пел- ликулярных гребней, которые при этом теряют свои правильные очертания и сглаживаются. Поверх упоминавшейся'-выше «базальной мембраны» (с. 143), которая не меняет своей формы, образуется своеобразная «присоска». Последняя плотно прикрепляется к гребням заднего конца тела примята. Образующиеся при этом замкнутые пространства заполняются многочисленными включениями, природа которых пока не известна.
же еще более тесного взаимодействия грегарип в сизигии может служить обитающая в полнхетах Diplauxia ІіиШ. У этого вида пелликула в зоне контакта двух особей постепенно резорбируется (рис. 81, В), так что между последними возникают широкие цитоплазматические «мостики». По ним из одной особи в другую проникают каналы эндоретикулюма, что говорит о глубокой физиологической интеграции организмов.
. • Иіітсграцни;Особсіі, иходящі!Х- в состав; сизигия, иногда заходит очень далеко. В первую очередь это, гіо-видимому, наблюдается у тех видов, сизигии которых образуются молодыми гамонтами. У паразита поденок Enterocystis \ungoides, например, происходит слияние зоны пеллнкулярных гребней двух особен, так что пелликула одной пепоередстиешю переходит и пелликулу другой (рис. 81, В). Базальная мембрана паразитов и подлежащие слои эктоплазмы, однако, и это объединение не включаются. Примером
Следующий этап полового процесса —это формирование гаметоцисты. Образующие сизигий особи округляются, причем каждая из них принимает форму полусферы. У видов, тело которых разделено на прото- и деіітомернт, септа между этими двумя отделами исчезает. На поверхности сизигия выделяется оболочка, имеющая довольно сложное строение и состоящая из нескольких слоев, самый наружный из которых часто имеет слизистую природу.
Гаметогенез всегда начинается с митотического деления ядер гамоитов. В большинстве случаев этот процесс многократно повторяется, так что число дочерних ядер может быть очень велико Процесс дифференциации последних приводит к образованию ядер двух типов; соматических и генеративных. Первых сравнительно немного[6]; они характеризуются крупными-' размерами и и дальнейшем сохраняются в остаточном теле. Вторые представляют собой ядра будущих гамет; они значительно более многочисленными обладают небольшими размерами. Вокруг генеративных ядер обособляются участки плотной цитоплазмы (рис. 82, Л, В), которые в дальнейшем отпочковываются от гамоитов. Таким образом завершается формирование гамет. Неиспользованная цитоплазме гамоитов, содержащая соматические ядра, превращается в первичные остаточные тела. Количество гамет в гаметоцпсте у грегарип как правило, весьма значительно. Исключение составляют лиші некоторые виды, относящиеся к отр. Neogregarinida (Ophryocystis Mattesia и др.), у которых вторично происходит уменьшение числе гамет, формирующихся из каждой особи, до 1—2. Это, очевидно связано с наличием в их жизненном цикле шизогонии, соответственно компенсирующем"! столь низкую половую «продуктивность»
Важной и характерной особенностью полового процесса грега рин, отличающей их от Coccidiomorpha, является образование равного числа гамет из каждого гамонта, входящего в состав сизигия Объясняется это в первую очередь тем, что половой процесс гре- гарин осуществляется в замкнутом и сравнительно небольшом объеме гамстоцнсты, где практически все гаметы имеют рсалыіук возможность копулировать.Копуляция грегаріш может быть как изогамію», так и аніізо- гамной. В типичной форме изогамная копуляция встречается у сравнительно небольшого числа видов и чаще всего у Neogregari- nida (Ophryocystis, Mattesia, Lipocystis и др.). Анизогамия распространена значительно более широко, причем наблюдается весь переход от случаев, когда она едва заметна (рис. 82, Г), до появ- І
Рнс. 82. Образование гамет и зрелые гаметы грегаріш (из км. Граеее). А - 'образованно мужских гамет ВІуІіктрІшІііх Iongicullix-,
В — образованно женских гамет Stylocepluilus tongicullis; В — анн- зогамная копуляция Stylocephalus longicollis; Г—гаметы Мопо- cystis sp.; Д — гаметы Gonospora tesliculi:
і — соматическое ядро, 2 — генератишюе ядро, 4—— последовательные стадии обособления мужских гамет, 7— зрелая мужская гамотп, л—/0— мослсдоиатслыіыс стадии обособления женских гамет, // — зрелая женская гамета, /2 —зиготастиямн, расположенными на полюсах и закрытыми специальными' пробочками. Через эти отверстия и осуществляется выход спорозоитов в кишечнике животного хозяина, проглотившего зрелые ооцисты. У некоторых видов грегарин один из полюсов ооцнсты несет один пли два тонких нитевидных выроста — фила.мента.
Внутри ооцнсты начинается образование спорозоитов. Как и у всех споровиков, у грегарин имеет место знготпческая редукция: первое метагамное деление ядра зиготы — менотичсское. У большинства видов далее следует еще два деления, в результате которых образуется 8 дочерних гаплоидных ядер. Вокруг них обособляются участки цитоплазмы и формируются спорозоиты. Как уже отмечалось выше, споры, внутри которых располагаются спорозоиты кокцидий, у грегарин отсутствуют.

Рис. 83. Выведенію зрелых оошіет из гаметоциеты (из mi. Грасее). Л — Тгісіічгііі/іісіііі.ч piilchcr. некрите гаметоциеты по зкікіторічин.іюму піну; Б — Gregarina munieri гаметоцнета с иыиерпутыміі спородуктамн:
/ —сгонка, гаметоциеты; 2 — остаточное тело. З — ооцн- ста, 4 — вывернутые спородукты, 5 — цепочки підходящих ооцнет
У большинства грегарин ооциста содержит восемь спорозоитов. Однако имеются виды, у которых количество спорозоитов в ооцне- те .сокращается до четырех (Selenidi- um) или, наоборот, возрастает до нескольких десятков (Meroselenidium).
Спорогония у грегарин чаще всего протекает во внешней среде, куда в большинстве случаев выводи тся га методиста, заполненная молодыми ооцистами.
Освобождение последних из гаметоцп- сты после завершения формирования спорозоитов может осуществляться нескольки ми различными способами. Почти всегда, однако, важную роль і этом процессе играет остаточное тело гамстоцпсты, которое возни
каст и результате слияния первичных остаточных тел гамоптов.
В наиболее простых случаях при повышении влажности окру жающей среды остаточное тело быстро набухает п увеличиваете;
в размерах. В результате давление внутри гаметоциеты возрастает стенки ее лопаются, и зрелые ооцисты разлетаются в разные cTq роны (рис. 83, Л). j
У многих специализированных форм, в частности у роді Gregarina, за счет остаточного тела образуется дополнительны! слой, прилегающий к стенке гаметоциеты. На нем, в свою очередь формируется несколько слепозамкнутых и направленных внутрь трубочек — спородуктов. Набухание дополнительного слоя или его резкое сокращение приводит к увеличению внутреннего давления в гаметоцисте. При этом спородукты выворачиваются наружу наподобие пальца перчатки, пронизывают стенки гаметоцпсты и прорываются па своем свободном конце. Созревшие ооцисты со спорозоитами выбрасываются по спородуктам во внешнюю среду в виде длинных цепочек (рис. 83, Б).
Спорозоиты. Спорозоиты грегарин по своему строению очень близки к спорозоитам остальных Sporo/.oa. Снаружи они одеты

Рис. 84. Шизонты Ophry- ocystis hagenmiVleri п малі.шіпюііом сосуде жука (но Ложе):
1 — стейка мальпигисоого сосуда, 2 — ядра шизонтов
пелликулой, подстилаемой слоем микротрубочек. Передний конец тела песет коноид, преконоидалыіые кольца и родтрии. На боковых поверхностях тела располагается несколько микропор. Отличие же заключается в том, что у некоторых видов имеется обширная мукропалыіая вакуоль, проток которой проходит внутри коноида. О функциях этого образования у спорозоитов пока ничего не известно.
Как уже отмечалось выше, у подавляющего большинства грегарин спорозоиты превращаются в гамонтов п только в относительно немногочисленных случаях они дают начало шизонтам.
Шизогония и мерозоиты. Облигатная шизогония имеет место лишь у представителей отряда Neogregarinida. Данные о широком распространении шизогонии в отряде Archigregarinida в последние го-, ды не подтвердились: бесполое размножение, по-видимому, сохранилось лишь у единичных видов згой группы.
Шизонты грегарин могут локализоваться как внутри, так и внеклеточио в полости кишечника пли мальпигиевых сосудов. Форма и размеры их тола у разных видов варьируют в очень широких пределах (рис. 72, 84). По своему тонкому строению шизонты неогрегарин очень близки, к шизонтам других споровиков, хотя и обладают некоторыми признаками, которые встречаются только у грегарин. Одним из таких наиболее важных признаков можно считать сохранение у шизоптов мукроііа, сходного с мукроном примитивных Eugregarinidu. Мукропалыіая вакуоль, так же как и у последних, содержит топкие фибриллы. Преконоидалыіые кольца сохраняются, тогка как коноид во время превращения спорозоита в шизонт резорбируется. Питание шизонтов, по-вндимому, осуществляется через многочисленные микропоры, расположенные на поверхности их тела.
Образование мерозонтов идет путем шизогонии. Сами мерозои* ты, обладая типичным для. этой стадии строением, на своем перед» нем конце (так же, как и спорозоиты) несут мукропалыіую вакуоль. Последующее развитие мерозоитов может быть двояким. Одни снова дают начало бесполым поколениям, другие же превращаются в гамонтов.
Взаимоотношении грегарин с животными-хоз^евами
Несмотря па то что грегарнпы являются очень широко распространенными паразитами различных бссиозшшочиых животных, вопрос о характере отношений между ними н их хозяевами до сих пор нс имеет однозначного ответа. Большинство авторов склонны рассматривать грегарин как комменсалов, не вызывающих никаких патологических изменений у животиых-хозяев. В подтверждение приводятся данные о нормальном развитии насекомых (в частности, саранчи) даже в тех случаях, когда интенсивность заражения грегаринами была исключительно велика. Простейшие целиком заполняли кишечник насекомого-хозяпна и по сути дела должны были бы полностью блокировать его.
Неоднократно высказывалось мнение и о мутуалнстнческом характере отношений в рассматриваемой'системе «паразит — хозяин». Американская исследовательница Р. Самнер (R. Sumner), работавшая с «мучными червями» (личинки жука Tcncbrio molitor), пришла к выводу, что присутствие грегарин необходимо для нормального роста и способствует увеличению продолжительности жизни их хозяев. Последние, по ее предположению, используют витамины и ферменты, вырабатываемые простейшими. Эта гипотеза нашла поддержку у ряда паразитологов.
Анализ полученных Самнер результатов показал, однако, что они не могут считаться достоверными. Было установлено, что грегарины при определенных условиях оказываются весьма патогенными для своих хозяев. В тщательно поставленных опытах с теми же «мучными червями» любое отклонение условий, в которых содержались насекомые, от оптимальных приводило к гибели и задержке развития в первую очередь зараженных особей.
Таким образом грегарин, по-видимому, необходимо рассматривать как очень специализированных н древних паразитов. Их отношения с хозяевами в результате длительной совместной эволюции характеризуются отсутствием внешних проявлении антагонизма. Однако любые достаточно серьезные изменения физиологического состояния жнвотного-хозяииа, которые могут быть вызваны самыми различными причинами (голодание, изменение температуры окружающей среды и т. п.), приводят к нарушению этого неустойчивого равновесия.
Дополнение к классу Sporozoa
Еще по теме . ПОДКЛАСС GREGARIN1NA DUFOUR — ГРЕГАРИНЫ:
- ПОДКЛАСС OPALININA
- ПОДКЛАСС COCCIDIOMORPH А DOFFLEIN
- ПОДКЛАСС RHIZOPODA SIEBOLD — КОРНЕНОЖКИ
- ПОДКЛАСС AWXOSI'OKIDIA
- ПОДКЛАСС PHYTOMASTIGINA CALKINS, 1909
- ПОДКЛАСС ZOOMASTIG1NA CALKINS, 1909 ОТРЯД K1NETOPLAST1DA HON1GBERG, 1963
- ПЕРВЫЙ СЛОВОИЗМЕНИТЕЛЬНЫЙ КЛАСС
- ДЕВЯТЫЙ СЛОВОИЗМЕНИТЕЛЬНЫЙ КЛАСС
- КЛАСС PLASMOSPORIDIA SPRAGUE
- ЛИНЕЙНЫЕ ПРЕОБРАЗОВАНИЯ ГЛАГОЛЬНОЙ ОСНОВЫ В ЕЕ ФИНАЛЬНОЙ ЧАСТИ. Общие замечания