ОТРЯД PIROPLASMIDA WENXON, 1920,- І ПИРОПЛАЗМИДЫ
Отряд Piroplasmida — очень своеобразная группа простейших, систематическое положение которых и родственные связи с другими Protozoa долгое время оставались неясными.
Все пнроплазімпды — внутриклеточные паразиты.
Их развитие протекает со сменой хозяев. В позвоночных животных паразиты поселяются в эритроцитах п в клетках ретикуло-эпдотелиальной системы. У переносчиков, которыми чаще всего бывают иксодовые или аргазовые клещи, они поражают клетки различных внутренних органов. Пироплазмиды, по-видимому, обладают только беспо- дым размножением. Достоверные данные о наличии в их жизненном цикле полового процесса отсутствуют.Впервые представители этой группы были обнаружены и 1888 г. В. Бабссом (V. Babes) в крови животных (крупного рогатого скота), страдающих гемогло- биноурнГшой лихорадкой.
Пять лет спустя в 1893 г. американским ученым Т. Смиту и Ф. Кнльборну (Т. Smith and F. Kilbourne) удалось проследить пути циркуляции и природе возбудителя так называемой техасской лихорадки крупного рога того скота — опасного заболевания, свирепствовавшего в те годы на территории США и наносившего огромный ущерб животноводству. Они установили, что передача паразитов от одного животного к другому осуществляется с помощью переносчиков — иксодовых клещей. В том же 1893 г. К. Старкович (С. Starcovici) показал, что кровепаразнты, описанные Бабесом, и организмы, вызывающие техасскую лихорадку, очень близки. Он предложил называть их и честь первооткрывателя бабе- зиями (р. Babesia — syn. Piroplasma). Позднее этот род вместе с несколькими другими вошел в состав сем. Babesiidac.
С самого начала изучения бабезиид в литературе оживленно дискутировался вопрос об их систематической принадлежности. Обитание в эритроцитах послужило причиной объединения этих простейших с кровяными споровиками. Некоторые авторы в своих работах описывали в цикле развития бабезий жгутиковые стадии н соответственно предлагали рассматривать последних как видоизмененных жгу: тиконосцев.
Амебоидное движение паразитов и наличие только бесполого способа размножения заставили ряд ученых сближать бабезиид с корненожками.Практически все исследователи связывали решение вопроса о положении этих кровепаразитов в системе простейших с вопросом о наличии у них полоиого процесса. Еще ц самом начале нашего столетня н ряді' работ указывалось, что в организме клеща-переносчпка у бабезий имеет место половой процесе, впоследствии многие паразитологи, среди которых видное место принадлежит Е. Деннису (Е. Dennis), подтверждали эти наблюдения. Сам Дсшшс, который описал изо- гамную копуляцию и образование оокішеты, обнаружил у обследованного им вида Babesia bigemina элементы жгутикового аппарата (кннетосиму и рпзоиласт). На этом осноаашш он высказал предположение, ч го бабезнн— /ю очень примитивные споровики, занимающие промежуточное положение между специализированными гсмоспорндиямн н жгутиконосцами.
В то же время многие исследователи не смогли обнаружить никаких признаков, подтверждающих реальное существование полоиого процесса а жизненном цикле бабезий. В этой связи большое значение имела работа немецких зоологов П. Регеидаица и Э. Рейхсмова (Р. Regendanz tind Е. Reiclienow), изучивших развитие Babesia canis. Они же первыми высказали предположение о родстве бабезиид с саркодовымн. Позднее сходные данные были получены и многими советскими паразитологами — Е. М. Хсйспным, Е. А. Муратовым, М. В. Крыловым н др. Сейчас уже большинство зоологов считают, что в цикле развития бабе- знид половой процесс отсутствует.
История изучения представителей другого семейства — сем. Theiicriidae, включаемого ныне в состав отр. Piropiasmida, аналогична истории изучения бабезиид. Первым в 1897 г. обнаружил теіілсрип в крови больных коров выдающийся немецкий биолог Р. Кох (R. Koch). Он примял их, однако, за бабезий. Через 6 лет, в 1903 г., им были найдены в клетках рстпкуло-эидотелналыіой системы зараженных животных крупные цитоплазматические включения, которые в дальнейшем получили название «коховских шаров» пли гранатных тел.
Кох также считал их одной из стадий развития бабезий. Лишь в 1907 г. было установлено, что эти паразиты отличаются от настоящих бабезий. Их выделили в самостоятельный род Theileria, а позднее было обосновано и отдельное семейство.Развитие тейлорнй в позвоночных животных исследовались многими авторами. Особых противоречий при этом нс возникало. Что же касается развития паразитов в клощах-нереиосчнках, то здесь, как и в случае с блбезиямн, мнения ученых разделились. Паразитологи Р. Гоидер (R. Guilder), Э. Коудри (Е. Cowdry), Э. Серджент (Ed. Scrgeiil) и другие описывали половой процесс, завершающийся образованием зиготы, и даже спорогонию. В то же время многие
исследователи, среди которых видное место принадлежит Э. Ренхеиову, не смогли обнаружит! каких признаков, подтверждавших наличие полоиого про
цесса в цикле развития теплерпп. 13 литературе первая точка зрения получила более широкое распространение, чем вторая. Однако за 40 с липшим лет, прошедших со времени опубликовании результатов исследования Серджента с соавторами, никаких серьезных доказательств в пользу того, что в цикле развития тенлерип половой процесс действительно имеет место, не появилось. Количество же работ, авторы которых приходят к выводу, что полового процесса у Theile- riidae нет, заметно возросло.
Отсутствие полового процесса у пнроплазмид долгое время служило главным и фактически единственным доводом против объединения этих кровепаразитов со споровиками. Положение резко изменилось после опубликования первых электронно-микроскопических работ. В результате этих исследований было выявлено удивительное сходство ультраструктуры пнроплазмид и типичных споровиков. Более того, некоторые особенности их организации весьма напоминают таковые кровяных споровиком (с. 114). Филогенетическая связь отр. Piroplasniida с классом Sporo/.oa признается большинством современных исследователей.
В настоящее время и состав отряда Piroplasniida включается' несколько семейств. Однако бесспорным можно признать лишь существование трех из них: сем.
Babesiidac Росію, 1913,— syn. Piroplasmidae Franca, 1910, сем. Theileriidae, Du Toit, 1918, и сем. Daclylosomidae Jakowska ct Nigrclli, 1955.Жизненный цикл пнроплазмид
Развитие паразитов в позвоночном животном j
В настоящее время ппроплазмнды описаны у представителей' всех классов позвоночных животных, однако наибольшее количество известных видов приурочено к млекопитающим. Наиболее подробно изучены циклы развития представителен двух семейств: Theileriidae и Piroplasmidae, на которых мы остановимся более подробно.
Инвазионным началом у пнроплазмид служат мелкие одно-; ядерные тельца, ппокулйруемые переносчиком-клещом в кровяное русло позвоночного животного. В литературе эти образования очень часто, хотя и не совсем точно, называются спорозонтами *. Их судьба у представителей разных семейств различна. Спорозои-j ты Theileriidae для дальнейшего развития проникают в клетки ре-; тикулоэидотелналыіой системы. Как правило, они локализуются в лимфатических узлах, печени, селезенке и других внутренних органах. Попав в клетку, паразит округляется, превращается в тро- фозопт и начинает расти. В дальнейшем путем шизогонии из трофо- зоита образуется большое количество одноядерных мерозоитов, которые вновь поражают клетки ретикуло-эпдотелпя. Относительно крупные одноядерные шнзонты, размеры которых могут достигать 10—20 мкм, и получили название коховских шаров или гранатных тел (рис. 85). После смены нескольких поколений подобных экзо- эритроцитариых шнзоитов вновь образующиеся мерозоиты тейле- рніі проникают в эритроциты животного хозяина.
В цикле развития бабезиид тканевые стадии, как считают большинство исследователей, полностью отсутствуют. Спорозоиты, по- [7] павшие в кровяное русло позвоночного, сразу же проникают в эритроциты.
Превращение спорозоитов и мерозоитов в трофозоиты у пироплазмид сопровождается известным упрощением строения. Этот процесс весьма сходен с аналогичным процессом у кровяных споровиков (с. ПО). Исчезают пелликулярные мембраны паразита и роптрии.
Трофозоит округляется (рис. 86, А). В цитоплазме на этой стадии хорошо выражены каналы шероховатого эндоретику-
Рис. 85. Схема цикла развития Theileria:
/ — трофозоит в клетке рстикуло-эндотелня, 2, 3 — последователь- ііые стадии формирования «гранатного тела», 4 — свободные меро* * зонты, 5 — зндоэрнтроцитарные стадии развития паразита, 6 — ме- розонты в просвете кишечника, 7 — шнзонты в эпителиальных клетках кишечника клеща, 8 — мерозонты в гемолимфе клеща. Р — мерозонты и шнзонты в клетках слюнных желез клеща, /0 — мерозонты в яйцеклетках клеща
люма. Типичные митохондрии и аппарат Гольджи у паразитов отсутствуют. Но зато так же, как и у ряда видов р. Babesia, имеются вакуолеподобные образования, ограниченные двумя мембранами и полимембранные концентрические структуры (с. 111).
Спорным является вопрос о способе питания трофозоитов, которые локализуются не внутри паразитофррной вакуоли, а непосредственно в плазме эритроцита. На обычных микроскопических препаратах молодые трофозоиты часто имеют вид кольца, сходного с соответствующей стадией развития трофозоита малярийного плазмодия. Обширные вакуоли, содержащие гемоглобин клетки хозяина, были описаны и в первых электронно-микроскопических работах (рис. 86, А). Образование вакуолей первоначально связывали с процессом заглатывания гемоглобина, а сами вакуоли рассмат- ривались как пищеварительные.
Позднее, однако, выяснилось, что эти образования представ' ляют собой лишь очень глубокие впячивания поверхностной мемб- раны паразита. Обособления этих впячиваний в замкнутые вакуоли никогда не происходит. Ряд авторов на основании детальных элект- ронно-мнкроскопических исследований приходят к выводу, что

Рис. 86. Строение эндоэритроцитарных стадий пироплазмид. А — молодой трофозоит Antlieinosoma garnhaini-, Б — трофозоит р. Babesia-, В — формирование мерозоитов Anthcmosoma garnhaini (по разным авторам):
/ — ядро, 2 дпоЛные мимбрлиные структуры.
3 — скопление микротрубочек, 4—- шероховатый эндоретнкулюм, 5 —• вакуолсиодобные впячивания поверхностной мем • браны паразита, 6 — полнмембранные структуры, 7 —трубчатое выпячивание трофо* зонта, 8 — пелликулярные мембраны, 9— роптрни, /0 — цитоплазма эритроцитаглавным способом поглощения пищевых веществ пироплазмидами является пиноцитоз. Образующиеся пиноцитозные вакуоли поступают в систему ограниченных мембранами трубочек, имеющихся в цитоплазме паразита. Последние часто связаны с упоминавшимися выше лолимембраннымн концентрическими структурами. Предполагается, что именно здесь завершается процесс переваривания проглоченной пищи.
Кроме того, у трофозоитов р. Babesia описаны особые трубчатые образования (рис. 86, Б), которые далеко выдаются из тела паразита и открываются на поверхности эритроцита. Одни исследователи рассматривают их как приспособление для заглатывания плазмы крови. Другие же приписывают им экскреторные функции. Во всяком случае неоднократно удавалось наблюдать, что через трубчатые uupocTU и плазму крови удаляются скопления мембран. Так пли иначе по способу питания трофозопты ппроплазмпд заметно отличаются от трофозонтов малярийного плазмодия. Это выражается, однако, не только в существовании разных способов поступления пищевых веществ в клетку паразита. Различен, по- видимому, и химизм процессов пищеварения. При переваривании гемоглобина ппроплазмпды никогда не образуют гемозонн, что очень характерно для подавляющего большинства споровиков.

Рис. 87. Эритроцитарные формы представителей сем. Babesiidae. А — Babesia Ы- gerrtina; Б — Babesia bovis\ В — Babesia diver gens-, Г — Nuitallia sp. (no Хейсину)
Размножение тротрозоптов в эритроцитах осуществляется только бесполым путем. Хотя у разных представителей пироплазмид оно приобретает различные формы, в его основе, по мнению ряда исследователей, всегда лежат различные типы шизогонии. У видов, относящихся к р. Babesia, тро- фозоит дает начало двум дочерним клеткам — мсрозоптам. Последние имеют грушевидную форму и располагаются в эритроците иод углом друг к другу,' образуя при этом довольно постоянную для каждого вида фигуру (рис. 87).
Долгое время считалось, что существуют'' три самостоятельных рода: Babesia (рис. 87, Б), Piroplasma (рис. 87, Л) и Babesiella (рис. 87, В), отличающиеся друг от друга размерами эритроцитарных Стадии и их положением в клетке животного хозяина. Сейчас псе виды, относившиеся рапсе к этим родам, включены н один род Babesia (Крылов, 1471).
Для р. Nuttallia, также относящегося к сем. Babcsiidae, при бесполом размножении характерно образование 4 мерозоптов, располагающихся в виде мальтийского креста (рис. 87, Г). Еще более отчетливо множественный характер шизогонии выражен у некоторых представителей сем. Dactylosomidae. У Anthemosoma, например, формируется 8 мерозоитов (см. рис. 86, В).
Деление ядра во время шизогонии ппроплазмпд осуществляется тем же способом, что н у кровяных споровиков. Ядро значительно увеличивается в размерах и принимает сложную лопастную форму. Одновременно наблюдается синтез ДНК, сопровождающийся эндо- митотическим увеличением числа хромосом. Выросты материнского ядра, содержащие необходимый генетический материал, заходят в формирующиеся почки II обособляются.
’ Полностью сформированные мерозоиты ппроплазмпд близки по строению к мерозоитам остальных Sporozoa (рис. 88). Они обладают хорошо развитыми роптрнямн, полярными кольцами, пелли- кулярными мембранами. У Theileria и Anthemosoma удалось обнаружить и типичную микропору. Как и у Haemosporidiidea, у мерозоитов пироплазмид отсутствует коноид. Многие виды из этой ста- дим, очевидно, утратили и микротрубочки (рис. 8G, З, 88, Б), столь характерные для мерозоитов и спорозоитов других споровиков. Зрелые мерозоиты заражают эритроциты, в которых повторяется весь цикл развития паразита.
У представителей сем. Babesiidae бесполое размножение в эритроцитах, по-внднмому, представляет собой главный способ увели-

Рис. 88. Мерозоиты пироплазмид (Порше-Хеннере и Вивье). А —Babesia cam's; Б—Thcileria parva:
1 — пелликулярные мембраны, 2 — микротрубочки, 3 — шероховатый эндо- ретнкулюм, 4 — ядро, 5 — двойные мембранные структуры, 6 — роптрии, 7 ~ полярное кольцоРис. 89. Экзоэритроак- тарная шизогония Babesia Ьіцетіпа и просвете капилляров (по Крылову)
чения числа паразитов в организме позвоночного животного. Чтс же касается тейлерппд, то у них наиболее интенсивное размножение осуществляется, как уже говорилось выше, в клетках ретнкулоэпдо телня. Способность же эритроцитарных форм к бесполому размножению скорее всего сильно ограничена. Лишь в очень незначительном количестве эритроцитов (у некоторых видов эта величина составляет 1% от общего числа инфицированных клеток) удается наблюдать крестообразные формы {см. рис. 85), сходные с мальтийскими крестами размножающихся нутталий.
В последние годы М. В. Крыловым было показано, что бабези- иды могут размножаться непосредственно в плазме крови. В капиллярах внутренних органов, хорошо снабжаемых кислородом (чаще всего в легких), паразиты образуют крупные шпзопты (рис. 89). Последние дают начало большому количеству одноядерных особен.
Передача паразитов от одного позвоночного животного к другому осуществляется, как уже говорилось выше, С помощью переносчиков. Инвазионными для последних являются эритроцитарные стадии развития пироплазмид.
Развитие паразитов в переносчике ^
Специфическими переносчиками пироплазмид в большинстве случаев служат клещи надсем. Ixodoidea, причем, главную роль, несомненно, играют собственно иксодовые клещи (сем. Ixodidae). Роль аргазовых клещей (сем. Argasidae) в распространении пироплазмид, по-видимому, весьма, ограничена. Перенос представителен сем. Dactylosomidae, паразитирующих в водных позвоночных. животных (рыбах и амфибиях), возможно, осуществляется паразитическими рачками р. Argulus (отр. Branchiura).
Развитие в организме клеща-переносчнка в настоящее время изучено только'у нескольких видов пироплазмид. Как уже отмечалось выше, данные эти весьма противоречивы. В последнее время, однако* ряд исследователей приходит к выводу, что в основе размножения паразитов в переносчике, так же как и в позвоночном хизипне, лежат разные типы шизогонии.
Освобождение пироплазмид из эритроцитов происходит в просвете кишечника клеща. Большинство авторов указывают, что значительная часть паразитов при этом погибает. Наиболее интенсивно размножаются в организме переносчика представители р. Babesia. Уже в просвете кишечника клеща удается обнаружить небольшие шпзопты (рис. 90). Каждый из них распадается па несколько (2—6) мерозоитов булавовидной формы, которые в свою очередь проникают в эпителиальные клетки кишечника и снова приступают к бесполому размножению. При этом формируются значительно более крупные шизонты. В результате асинхронного почкования они дают начало нескольким десяткам (до 50) одноядерных булавовидных особей (рис. 90). Последние либо снова поражают эпителиальные клетки, либо проникают в полость тела хозяина. В гемолимфе паразиты сохраняют способность к размножению,'однако, процесс этот идет менее интенсивно. Из гемолимфы булавовидные формы паразитов проникают в различные внутренние органы: слюнные Железы, гонады, гиподерму и т. д. Размножение бабезий в клетках слюнных желез, также протекающее путем шизогонии (рис. 90), приводит к образованию мелких грушевидных мерозоитов, которые инвазионны для позвоночного животного. Они иесколь-
ко отличаются от аналогичных стадий, образующихся при агамном размножении паразита в других органах клеща, как морфологически, так и физиологически. Грушевидные мерозоиты почти в два раза мельче булавовидных, а главное они физиологически «преадап- тированы» к обитанию в кровяном русле позвоночного животного.
13 гонадах паразиты прежде всего поражают яйцеклетки, которые при этом не теряют способности к нормальному развитию. Таким образом осуществляется трансоварналыіая передача бабе-

Рис. 90. Pxmimic linbcsia Ыцстіпа и клсщс-исрспосчико (по Муратову и Хейсипу):
/ — ппгіомт ції просища кишечника клеща, 2 — ішіаоит на эпителиальных клеток кишечника клеща, 3 — булавовидные мерозоиты из гемолимфы, 4— шмзоит и грушевидные мерозоиты из слюнных желез клеща
знй. Сформировавшиеся из таких зараженных яиц клещи несут в клетках своих внутренних органов, в том числе и в слюнных железах, большое количество паразитов. При нападении на позвоночных животных для питания они инокулируют в его кровяное русло инвазионные мерозоиты.
Развитие в переносчиках представителен р. Nitllallia протекает в основном сходным образом.
Однако трапсоиарпалыюй передачи у них обнаружить не удалось.
Сведения о развитии тейлерий в переносчике очень противоречивы. Большинство исследователей сходится на том, что паразиты размножаются в слюнных железах клеща (во многих работах этот процесс рассматривается как спорогония). По-видимому, паразиты способны размножаться и в просвете кишечника и в его эпители-
алыіых клетках, хотя її то и другое некоторыми шпорами оспаривается. Инвазируют они и другие внутренние органы клещей.
Очень спорен вопрос и о возможности трапсоварпальнон передачи тейлерий. Долгое время большинством исследователей принималось, что у этой группы пироплазмнд передача простейших от одного поколения переносчиков к другому отсутствует. В последние годы, однако, появился ряд работ, в которых экспериментально показано существование такого пути циркуляции паразитов. Вероятно, что трансовариальная передача свойственна лишь отдельным видам тейлерий, а может быть, даже и отдельным штаммам.
Заболевания, вызываемые иироплазмлдалш
К отр. Piroplasmida относятся возбудители очень серьезных и опасных заболевании домашних животных. В соответствии с родовыми названиями пироплазмид они обозначаются как баоезпозы (пироплазмозы), нутталиозы и тейлериозы. Хотя в клиническом проявлении этих заболеваний имеется некоторое сходство, каждое из них характеризуется своими диагностическими признаками. От бабезиозов (возбудители: Babesia bigernimi, В. bovis, В. ovis и др.) и теіілернозов (возбудители: Tlicileria аппиіаіи. Т. рагѵа. Т. inulans и др.) страдает в основном крупный и мелкий рогатый скот; нутталлии же (Nuttallia equi) главным образом поражают представителей сем. Equidae — лошадей, ослов и т. п.
Все эти заболевания наносят огромный ущерб животноводству. Особенно широко они распространены в районах с теплым климатом, где часто принимают характер опустошительных эпизоотий. Экономические потери обуславливаются не только падежом скота, но и резким снижением его продуктивности. Например, у коров, переболевших телериозом, надои восстанавливаются лишь на 70-75%.
Клинические признаки проявляются не сразу после ппикулнцпи паразитов в кровяное русло животного, а через определенный промежуток времени. Продолжительность инкубационного периода в среднем составляет 10—15 дней. Заболевание сопровождается сильным подъемом температуры, увеличением внутренних органов— печени, селезенки. При заражении тейлериями набухают лимфатические узлы. Развішается анемия. Последняя усиливается благодаря гемолизу эритроцитов. При бабезиозах, часто называемых «кровемочкой», это сопровождается отчетливо выраженной гемо- глобиноурией, которая проявляется уже на 2—3-й день после заражения животного. Для теіілернозов характерно заметное повышение проницаемости кровеносных сосудов, что приводит к возникновению многочисленных кровоизлияний как во внутренних органах, так и па открытых участках кожи п слизистых. Серьезные морфологические и функциональные нарушения наблюдаются и пищеварительной, выделительной и нервной системах. Все это вызывает ослабление больного животного, угнетение, потерю аппетита.
Сильное истощение и кишечная атрофия приводят к потере подвижности. При отсутствии лечения больные животные часто погибают.
При спонтанном выздоравливании животных, когда всякие клинические проявления заболевания исчезают, полного освобождения организма хозяина от паразитов не происходит. Простейшие еще долго остаются в кровяном русле. Именно такое паразптопосптель- ство, по мнению многих авторов, является причиной невосприимчивости животных к повторным заражениям. В районах, неблагополучных по заболеваниям, вызываемым пироплазмндамн, страдает привозной скот. Местные же животные в основном переболевают
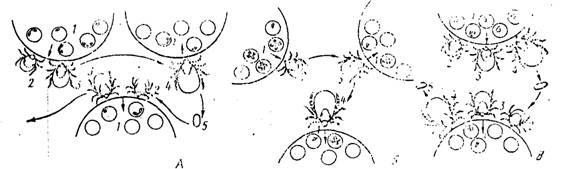
Рш-. 01. Пути ж'|и'дачі! ііпріж.'іазміїд or одного по.чііоно'іііого нанигі нот к другому и природо. Л -- - передача Оабоаіііі клещами о дііуххішішіиим циклом разніі- чия (Rlupiccpludus)-, Б — передача бабезиії клещами с одііохозяшшым циклом развития (Boophilus); В — трансфазная передача нутталлнГі клещами с трех- хозянииым циклом развития (Dermacenior);
/ — эритроцитарные стадии развития паразитоп, 2 — личинка, 3 — нимфа, •/ — имаго,
5 — яПцо
еще в молодом возрасте. Возможность длительного сохранения паразитов в кровяном русле и последующая ревакцинация при повторных нападениях иивазированных клещей поддерживают устойчивый иммунитет, который не дает развиваться тяжелому заболеванию.
Характер развития ппроплазмнд в переносчиках определяет и некоторые особенности природных очагов заболеваний. Представители р. Babesia, для которых свойственна трансовариальная передача паразитов от одного поколения клещей к другому, могут сохраняться в природе длительное время даже при отсутствии позвоночных животных. В литературе имеются данные о сохранении клещами паразитов на протяжении 50 поколений. Таким образом, даже прекращение выпаса скота в том или ином опасном районе не приводит к исчезновению возбудителей. Наличие трансона- рналыюй передачи, кроме того, позволяет паразитам использовать в качестве переносчиков клещей как с двух- и треххозяннпымп циклами развития (виды |>. Rliipicei>li(ilits, llaentophysalis, Ixodes и др.), так и виды, нее развитие которых от личинки до имаго осуществляется только на одном животном хозяине. В первом случае, если личинки іі нимфы питаются на животных того же вида,к..
что и имаго, передача паразитов может осуществляться трніісфазпо (рис. 91, А). Неполовозрелые стадии развития клещей заражаются пироплазмндами при иападснин на больное жпг.отпое. После линьки половозрелая фаза имаго уже способна заражать новых хозяев. Во всяком случае, при одпохозяпппом цнк.'іе развития переносчика (виды р. Boophilus) заражение новых позвоночных животных осуществляется клещами следующего поколения (рис. 91, В).
Нутталлии и, по-видимому, большинство тендернії, у которых отсутствует трансовариальная передача, способны использовать в качестве переносчиков лишь клещей с двух- її треххозяпппыміі циклами развития (Вепіинеп/ог, 11 i/uh пиши, / / ui’iiinplitjsul is. Р/іірі- cepliulus, Ixodes). У них передача паразитов протекает только трансфазно (рис. 91, В).
Борьба с заболеваниями, вызываемыми лнроплазмпдамп, cof^" ит из комплекса мероприятии. Специфические хпмиопрепараты, воздействующие на паразитов в позвоночном животном, отсутствуют. В различных сочетаниях с антибиотиками, витаминами, кофеином, глюкозой и пр. применяются такие препараты, как акаприн, аминоакрихнн, гемоспоридин, бнгумаль, берспил и др.
Трудность лечения этих заболевании резко повышает значение профилактических мероприятий, большинство из которых основано на создании условий, исключающих возможность контакта клещей - и домашних животных. Создание культурных пастбищ и загонный выпас скота на них, стойловое содержание молодняка, использование акарицидных препаратов для уничтожения клещей, питающихся на животных, и для обработки помещений, в которых содержится скот,— все это позволяет добиться резкого снижения заболеваемости. IJ последние годы нее шире и шире используется метод химиопрофилактики кровепаразитарных заболеваний скота. Важное значение приобретает и метод иммунизации скота тканевыми культурами паразитов.
Положение пироилазмид в системе простейших
Относительно положения пироплазмид в системе Protozoa до сих пор нет единого мнения. Наличие филогенетической связи между пироплазмидами и споровиками подвергалось сомнению многими авторами. Основным доводом против включения пироилазмид в класс Sporozoa было и остается отсутствие у первых полового процесса. Вместе с тем сходство в организации обеих групп так велико, что объяснить его лишь конвергенцией не представляется возможным. Существует ряд внутриклеточных паразитов; однако никто из них под влиянием образа жизни не приобрел сложно устроенного апикального комплекса органоидов, столь характерного для споровиков. Нс встречается в других группах и своеобразный способ бесполого размножения шизогония. Вес ли подтверждает точку зрения тех исследователей, которые рассматривают пироилазмид как настоящих Sporozoa, утративших, однако, половой процесс и обладающих абберантным жизненным циклом.
О том, что такая возможность реально существует, говорит пример токсоплазм и саркоспорпдин (с. 97), которые могут неограниченно долгое время наразитировать только в промежуточных хозяевах, размножаясь агампым путем.
Из всех представителен класса Sporozoa иироплазмпды, по-шіди- мому, ближе стоят к кровяным споровикам. Это подтверждается общими чертами строения и особенностями деления ядра (с. 106). В то же время они обладают и рядом очень своеобразных призма* ков, говорящих о еще более глубокой спеішалнзаціш, чем что имеет место у кровяных споровиков. Наблюдается дальнейшее упрощение морфологии расселптельны.х п инвазионных стадпіі - - меро- зоптов. Паразиты локализуются непосредственно в строме эритроцита, а не в паразптофорпоп вакуоли. Они обладают иным типом переваривания гемоглобина, и, наконец, самое главное, у них исчезает половой процесс. Все это не позволяет объединять пироплазмид с какой-либо другой известной группой Sporozoa. Их приходится рассматривать как. самостоятельный отряд подкласса Coccidiomorpha.. Piroplasmida, по-вндпмому, имели общих предков с кровяными споровиками.
Основные направления эволюции споровиков
Споровшем — цельная п единая по своему происхождению группа паразитических простейших. Для них характерна однотипность своеобразных морфологических структур и способов бесполого размножения. Единым, несмотря на многочисленные вариации и отклонения, представляется и ход жизненного цикла споровиков, включающий чередование бесполого размножения (шизогонии), полового процесса и спорогонии.
Эволюция жизненного цикла споровиков осуществляется по трем основным направлениям. Это вторичное сокращение хода жизненного цикла, его вторичное же усложнение п изменение соотношений эндогенной п экзогенной частей цикла.
В отдельных случаях ход жизненного цикла упрощается за счет выпадения стадий бесполого размножения (отр. Eugregarinida, отр. Protococcidiida). Высокая плодовитость паразитов, необходимая для существования вида, поддерживается при этом за счет появления «компенсаторных механизмов», обеспечивающих увеличение числа формирующихся ооцист, например образование сизи- гиев и т. п. (с. 147).
Усложнение жизненного цикла споровиков происходит за счет включения промежуточного хозяина. Появление последнего может быть обусловлено особенностями биологии дефііііптпіпіого хозяина, іілирі!'що і-.-ѵ '.ншншдѵпчп: Дей';!.1.-;:гшшк’Г; хозчнн -- хищник — хе
Н.чссГ ЛЛJ О Г 0 Л ОН ГОЛ Г- ѵ Л0'-І50Л ЛЛЛ ЛрЛЛСНЛЬЛМ Л ЛОМ {\ БОЛ.ЧЗЛЛ
споровиков), где обычно сосредотачиваются выведенные наружу ооцисты. Это затрудняет возможность его заражения. Передача инвазии осуществляется в таких случаях по пищевой цепи в системе хищник — жертва. Роль же промежуточного хозяина исполняют животные, непосредственно контактирующие с грунтом (например, грызуны, травоядные животные н т. п.). Жизненные циклы такого типа свойственны р. Sarcocystis и р. Toxoplasma (с. 97), морским кокцидпям Aggregalct (с. 86) и т. д. В других случаях роль промежуточного хозяина могут выполнять различные кровососущие беспозвоночные (пиявки, клещи, насекомые), передающие паразитов от одного позвоночного животного к другому. У примитивных видов (сем. Lankeslerellidae, с. 86) это, в сущности, еще переносчик, в котором паразиты не претерпевают никакого развития. В то же время у более специализированных групп (итр. Adeicida, отр. Piroplasmida, подотр. Haemosporidiidea) в промежуточном хозяине протекает значительная часть жизненного цикла (гаметогенез, копуляция, спорогония).
Для большинства споровиков характерно чередование эндогенной и экзогенной.частей цикла (с. 84). Однако, как и у других паразитов, у споровиков отчетливо проявляется установленная В. Л. Догелем тенденция ко вторичному сокращению тех стадий, развитие которых осуществляется во внешней среде.
По-видимому, наиболее примитивным в этом отношении следует считать жизненный цикл р. Elvtillwrosclii’oit (отр. Prolococcidiida), у которого в кишечнике хозяина (полпхеты) осуществляется только шизогония и развиваются гамопты. Последние выводятся во внешнюю среду, где и протекает формирование гамет, копуляция и спорогония. У всех же остальных споровиков половой процесс, как правило, смещается на период пребывания паразитов в теле хозяина. У некоторых кокцидип это смещение идет еще дальше и даже ооцисты полностью созревают в организме хозяина и выводятся наружу инвазионными, т. е. содержащими спорозоитов (р. Sarcocystis, р. Aggrcgala и др.).* Длительность экзогенной стадии при этом резко сокращается. Наконец, в ряде случаен экзогенная часть цикла полностью выпадает, и передача инвазионных стадии от одной особи хозяина к другой осуществляется при помощи переносчика-кровососа. Типичным примером может служить жизненный цикл малярийного плазмодия.
Предки современных споровиков, по-впдпмому, были паразитами беспозвоночных животных, скорее всего каких-то первичных Coelomata, от которых в дальнейшем произошли кольчатые черви и членистоногие. Характерно, что именно у кольчатых червей паразитируют наиболее примитивные представители Sporozoa (Archigre- garinida, Protococcidiida и др.). Почти исключительно с членистоногими связана и эволюция грегарпн. Coccidiomorpha тоже довольно широко представлены в членистоногих, но становление этой группы, по-видимому, совершалось преимущественно в позвоночных животных.
Большая часть споровиков — внутриклеточные паразиты, т. е. специализированные организмы, способные существовать лишь при строго определенных условиях. Такая узкая специализация вряд ли может быть первичной. Предки современных споровиков, вероятно, были полостными паразитами, локализовавшимися скорее всего в целоме или в просвете кишечника хозяина. Такая локализация сохранилась у представителей примитивного отряда Protococcidiida паразитирующих в морских беспозвоночных.
Переход споровиков к внутриклеточному паразитизму, несомненно, результат постепенной и длительной эволюции. О ее последовательных этапах до известной степени можно судить по аберрант ному ходу жизненных циклов некоторых современных видов, по ха рактеру миграции паразитов в организме хозяина и т. п. Особыі интерес в этом плане представляет жизненный цикл Toxoplasmc gondii и некоторых видов р. Еітегіа. Высвободившиеся из ооцпсть спорозоиты этих видов не задерживаются в кишечнике хозяин; п, минуя кшпечиый эпителий, мигрируют в субмукозу, где и захва тывнются макрофагами. Дальнейшая судьба спорозоитов различна Подавляющее большинство их, покинув через некоторое'время мак рофаги, возвращается обратно в кишечник и поселяется в клетка: кишечного эпителия. Незначительная же часть внедряется в клетю внутренних органов, где превращается в трофозонтов іі дает начал» ряду сменяющих друг друга агамных поколений. Спорозоить Т. gondii, попавшие в организм промежуточного хозяина (грызуні и др.), вообще не возвращаются в кишечник, оставаясь впутрпкле точными паразитами различных органов (с. 97).
І Ін тересно, что участие макро(|>агов в качестве клеток-хозяеі па первом этапе^жпзнепного цикла описано не только для спорови ков, но п для других простейших. Например, для микроспорндиі (с. 183) и лейшмашій. Это нельзя рассматривать как случайно совпадение. Вероятнее всего во всех перечисленных случая, мы имеем дело с рекапитуляцией одного из начальных этапов пбре хода простейших к внутриклеточному паразитизму. Вполне естест веппо ожидать, что филогенетически первыми «хозяевами» для ни. должны были стать именно клетки-фагоциты, активно захватыва ющпе все инородные тела, попадающие в микроорганизм. Процес же адаптации простейших к внутриклеточному паразитизму, оче видно, шел по нуги преодоления защитной реакции макрофаго и использования их в качестве среды обитания.
Переход споровиков от поселения в полости тела к впутрикле точному паразитизму мог осуществляться в результате случайног попадания спорозоитов в кишечник животного хозяина. Всроятж паразиты стремились уйти из кишечника, как из среды иеблагс приятной («реакция бегства», по определению В. Догеля. Оказаі шись за пределами кишечной полости, в лимфе или ретикуло-эндс телиальной системе, они неизбежно должны были стать «жертвами макрофагов. Большая часть паразитов при этом, конечно, уннчте жалась, по немногие выжившие могли дать начало ряду поколенії! постепенно адаптировавшихся к жизни и цитоплазме макрофагоі Отсюда нетрудно представить и переход паразитов к поселенні в клетках других типов. Первоначально паразиты, вероятно, пре пикали в самые различные клетки. Такая «неразборчивость» сохра лилась и у некоторых современных споровиков — Toxoplasma gondii
Lankesterella minima и др. Это, несомненно, очень прпмпа пішая черта названных видов.
Дальнейшая эволюции предкових форм Spurozoa, по-іш,німому, шла но пути специализации и закреплении способности паразитировать в клетках лишь строго определенного типа. Мы зпае.м виды, избирательно поселяющиеся в саркоплазме мускульных клеток {Sarcocystis), в эндотелии сосудов (Hacmoprotcits и др.), в форменных элементах крови (все кровяные споровики, некоторые Adeleicla и др.), в эпителии почечных канальцев (Кітегіа Іпшсаіа) и, наконец, в клетках кишечного эпителия (подавляющее большинство кокцнднй).
Приуроченность споровиков к клеткам тех или иных тканей позвоночного связана с проблемой расселения вида. Для параш: которые локализуются во внутренних органах, изолированных от внешней среды, расселение может иметь место только при поедании одного хозяина другим. В то же время локализация паразитов в органах, связанных с внешней средой (например, в кишечнике), обеспечивает возможность свободного выхода наружу расселитель- ных стадий. Вероятно, именно поэтому паразитирование споровиков в кишечнике приобрело столь широкое распространение. Почти также часто встречаются споровики, паразитирующие в клетках' крови. В этом случае «проблема расселения» паразитов решается участием в их жизненном цикле кровососущих беспозвоночных — переносчиков. Биологическая роль последних в процессе осуществления цикла может быть различной у разных видов споровиков.
Анализ особенностей жизненного цикла современных кишечных кокцидий и грегарин помогает попять пути перехода споровиков к паразитированию в клетках кишечного эпителия хозяина. По-видимому, древние споровики, уже на самых ранних этапах их эволюции, были не просто паразитами кишечника, а вступали в тесную связь с клетками кишечного эпителия. На это указывает, например, тот факт, что трофозоиты архигрегарии питаются за счет эпителиальных клеток, погружая в них мукроп; то же характерно для примитивных представителей Eugrcgarinida. Гамопты, упоминавшегося уже р. Eleutheroschizon (отр. Protococcidiida), поселяются непосредственно на поверхности клеток кишечного эпителия Хозяина (полихет). Даже среди таких специализированных споровиков, как кокцидии сем. Eimeriidae, встречаются виды (Cryptosporidium), весь жизненный цикл которых проходит в области «щеточной каймы». Биологически подобная приуроченность паразитов к зоне микроворсинок легко объяснима. Именно здесь осуществляется пристеночное пищеварение хозяина, что обусловливает высокую концентрацию в этой зоне как пищевых веществ, так и ферментов.
Переход от жизни на поверхности клеток к внутриклеточному паразитированию, вероятно, осуществлялся постепенно. Сначала образовывалось глубокое впячиванне наружной мембраны клетки, в которое паразиты погружались лишь частично. Позднее они превратилось в замкнутую паразптофорпую вакуоль. Последовательные этапы этого процесса можно понять на примере хода жнзнеп- пых циклон никоторых современных кокцпдш'і р. Пітегіа. Показателен в этом птпшпсипп кпд Пітегіа ип^иШае (паразит угря). Трофбзопты этих кыщидпп живут п размножаются па поверхности эпителиальных клеток, п аопе «щеточной каі'імы». Там же проходит и формирование мпкрогамет. Ипаде расположены мпкрогаметы, которые па половину своего диаметра погружены в клетки кишечного эпителия. На поверхности последних образуются чашевидные впячпвания. После оплодотворения зигота целиком уходит в глубь эпителиальной клетки и превращается там в ооцпсту. ТІллюстра-! цпеіі следующего этапа перехода к внутриклеточному паразитизму может служить жизненный цикл Dobellia (отр. Aclclcicla), паразитирующей в кишечнике еппупкулпд. У этого вида толіжо шизогония осуществляется па поверхности эпителиальных клеток. Развитие же гамоптов, гаметогенез и формирование ооцпст протекает впутри- клеточпо. Непосредственные причины, обусловившие переход кн- піечных споровиков к внутриклеточному паразитизму, могли иметь двоякий .характер. Во-первых, частичное, а тем более полное погружение паразитов в глубь клеток исключает опасность быть вынесенными из кишечника во внешнюю среду. Во-вторых, существование внутри клеток хозяина позволяет простейшим наиболее полно использовать их для обеспечения собственных потребностей.
Переход споровиков к внутриклеточному паразитизму связан с глубокой специализацией ц вторичными изменениями морфологии и физиологии паразитов. Особенно отчетливо прослеживаются эти изменения у тех видов, которые приспособились к паразитированию в эритроцитах (1 lauiiosporidiidea, Р ігор I asm і cl а). і Іх спорозоиты и мерозопты утратили коноид, микротрубочки развиты у них значительно слабее, чем у других споровиков, у некоторых видов исчезают митохондрии и*т. д. Физиологическая специализация этих паразитов заходит столь далеко, что некоторые из них (например, малярийный плазмодий) теряют способность к осуществлению собственных обменных процессов и получают жизненно необходимые ферменты только из клеток хозяина (с. 112).
Подводя итоги, можно выделить несколько основных направлений, но которым шла эволюция споровиков.
1. Возникновение единого, характерного только для споровиков плана строения (в особенности для расселптелыіых и инвазионных стадий). 2. Переход от паразитирования в беспозвоночных животных к позвоночным. 3. Переход от обитания в полости тела или в просвете кишечника хозяина к внутриклеточному паразитизму. 4. Стабилизация жизненных циклов, которые в наиболее полном виде включают чередование половых и агамно размножающихся поколений. Отклонения от этой схемы, приводящие либо к упрощению цикла (выпадение бесполого размножения), либо к его усложнению (появление дополнительного бесполого размножения в промежуточных хозяевах) носят вторичный характер. 5. Сокращение сроков развития экзогенных расселптелыіых стадии во внешней среде и, в конечном счете, полное их выпадение в связи с появлением псреносчнков-кронососов.
Еще по теме ОТРЯД PIROPLASMIDA WENXON, 1920,- І ПИРОПЛАЗМИДЫ:
- ОТРЯД DIPTERA-ДВУКРЫЛЫЕ
- Отряд многожгутиковые (Polymastigina)
- Отряд Кокцидий (Coccidia)
- ОТРЯД POLYMASTIGIDA (BLOCHMANN) EMEND. REICHENOW
- У СТУДЕНТОВ ЕСТЬ СВОЯ ПЛАНЕТА... Студенческие строительные отряды
- ОТРЯД POLYMASTI01NA
- ОТРЯД ACARIFORMES-АКАРИФОРМНЫЕ КЛЕЩИ
- ОТРЯД HEMIPTERA (HETEROPTERA)- КЛОПЫ
- ОТРЯД ANOPLURA (S1PHUNCULATA) - ВШИ
- ОТРЯД BLA TTODEA- ТАРАКАНОВЫЕ
- ОТРЯД PERITRICHA STEIN-КРУГОРЕСНИЧНЫЕ
- ПОДКЛАСС ZOOMASTIG1NA CALKINS, 1909 ОТРЯД K1NETOPLAST1DA HON1GBERG, 1963
- ОТРЯД DII’LOMONADIDA WliNYON, 1926
- ОТРЯД PROTOMONADINA
- ОТРЯД HAEMOSPORIDIA
- ОТРЯД CYCLOPHYLLIDEA
- ОТРЯД AMOEBINA-АМЕБЫ
- ОТРЯД COCCIDIA- КОНЦИДИИ
- ОТРЯД pseudophyllidea-лентецы
- ОТРЯД APHANIPTERA - БЛОХИ