ПОДОТРЯДЫ OXYMONADINA (HONIGBERG, 1964) И HYPERMASTIGINA (GRASSI ЕТ FOA, 1911)
Чрезвычайно своеобразные и богатые представителями группы паразитических жгутиконосцев (рис. 30), особенности строения и биологии которых были раскрыты в результате исследований целого ряда известных ученых — протистологов (В.
А. Догель, L. Cleaveland, Н. Kirby, Р.' Grasse и др.). К этим подотрядам относятся крупные (до 0,5 мм длиной) жгутиконосцы, которые обитают в задней кишке термитов и питающихся древесиной тараканов Cryptoccrcus.Морфология. Для этих жгутиконосцев очень характерна тенденция к полимеризации тех или иных органоидов, чаще всего жгутиков и парабазальных тел (Hypermastigina), а в отдельных случаях и ядер (некоторые Oxymonadina). Особенно многочисленны бывают жгутики, которые в числе многих сотен, а иногда и тысяч располагаются на переднем конце тела (Barbulanympha), простираются до его середины (Trichonympha) или достигают заднего конца (Teratonympha). Крупное ядро в большинстве случаев
"I
Рис. 30. Разные жгутиконосцы из кишечника древоядных тараканов и термитов. Л — Lophomonas; Б — Leptospironympha- В — Іоепіа\ Г — Spirotrichonympha; Д — Snyderella (по Яницкому, Кирби и Кливленду)находится в передней трети тела. Парабазальные тела многочисленны. Лксостиль обычно хорошо развит у представителей Охуто- пасііпа. У Hypermasligina он иногда имеет вид полой трубки, в передней расширенной части которой помещается ядро (сем. Lopho- monadiilae). Органоид, играющий опорную роль, становится, таким образом, еще и защитным образованием для ядра. Но в ряде семейств аксостиль представлен отдельными фибриллами (сем. Spi-

Рис. 31. Строение Trichonympha collaris (no Кирби):
/ — шапочка, 2 хоботок, 3 — продольные эктоплазматические складки, 4 — ядро, 5 — парабазальные тела, G — жгутики
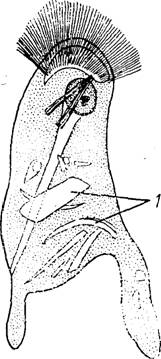
Рис.
32. Жгутиконосец Joe- nia intermedia в стадии образования цитоплазматических выростов для захвата пищевых частиц (по Догелю) :/ — захваченные частицы древесины из содержимого кишеч- ника хозяина
rotrichonymphidae) или совсем отсутствует (сем. Trichonymphidae). Передний конец тела нередко очень сложно дифференцирован. Так, у представителей сем. Trichonymphidae он преобразован в хоботок (рис. 31), отграниченный от тела глубокой перетяжкой. По центральной оси хоботка проходит тонкая трубка. В прилежащем к ней внутреннем слое эктоплазмы находятся многочисленные ки- нетосомы. Наружный слой эктоплазмы кажется ребристым из-за множества тонких, тесно сближенных продольных складок, расположение которых напоминает сложенные листы книги. Из щеле- вндных промежутков между ними выходят наружу ряды жгутиков. Вершина всего этого сложного образования прикрыта плоской шапочкой.Питание, размножение и жизненный цикл. Наиболее обстоя телыю изучены на примере Hypermasligina. У этих жгутиконосце нет цитостома. Тем не менее они питаются оформленными пище вымн частицами — кусочками древесины из содержимого кишеч пика хозяина. Процесс поглощения этих частиц подробно опнсаі Кливлендом. Кусочки древесины приклеиваются к заднему конц; тела жгутиконосца, где вслед за тем образуется впячйвание в виді глубокой чаши (рис. 32). Края чаши вскоре смыкаются, и пищевьп частицы оказываются заключенными внутри большой вакуоли, і которой и происходит их переваривание. Некоторые ITypermastigi па питаются подобно амебам, захватывая пищевые частицы с по мощью псевдоподий.
В период линьки термитов и древоядпых тараканов больша; часть жгутиконосцев выбрасывается из кишечника вместе с хитн новой выстилкой прямой кишки. Однако непосредственно послі линьки хозяина наблюдается вспышка размножения жгутикоирс _цев. Интересно, что в промежутках между линьками жгутиконосць почти не размножаются. Это один из примеров весьма распрост раненного явления — строгой приуроченности жизненного- ціщл; паразита к жизненному циклу хозяина (Догель, 1947, 1962).
Ну permastigina не могут существовать вне кишечника хозяина, но і хозяева не могут жить без своих паразитов. Как было установлене Кливлендом, дефаунированные, т. е. освобожденные от жгутико носцев, термиты быстро гибнут, так как не способны без их участіи переваривать клетчатку, которой питаются. Жгутиконосцы же вы рабатывают фермент целлюлазу, гидролизующий клетчатку Огромное количество жгутиконосцев (в кишечнике термитов н: насчитывают до 25 000!), «работающих» на хозяина, по-видимому вполне обеспечивает его потребности в пище. Вот почему в после лииочный период так важно быстрое восстановление численності жгутиконосцев. У термитов и тараканов эти процессы осуществил ются по-разному. Hypermastigina из тараканов способны, подобие огромному большинству простейших, образовывать цисты. Сам-ш процесс инцистирования стимулируется линочпыми гормонами хо зяина, и цисты обычно появляются в период, предшествующи!' линьке. Благодаря этому во внешнюю среду выбрасывается боль шое количество цист, сохраняющих жизнеспособность при небла гоприятных условиях. Перелинявшие тараканы и их личинки зано во заражаются необходимыми для них паразитами, заглатыва? рассеянные во внешней среде цисты. Иначе обстоит дело у терми тов. Паразитирующие в них виды Hypermastigina не способрь инцистироваться и, попадая во внешнюю среду, сразу же погибают Личинки термитов, только что вышедшие из яиц, или перелиняв шиє іГПОТомулпшенные жгутиконосцев, заражаются, слизывая ка пельки экскрементов, выступающие из анального отверстия другю термитов.В экскрементах всегда имеется некоторое количество жгутико носцев. Однако прежде чем попасть в прямую кишку заглотившей их термита, они должны пройти его желудок и средний отдел КП шечннка, где условия могут быть неблагоприятны н даже губи- тслыіы для них. В связи с этим у I Iypcrmasligina, паразитирующих в термитах, выработались особые приспособления, в известной мере заменяющие инцистирование. По наблюдениям В. А. Догеля, жгутиконосцы могут выталкивать из цитоплазмы все пищевые частицы, иногда даже целиком отделять заднюю часть тела.
Цитоплазма их при этом сжимается, жгутики сбиваются в пучки и покрывают все ссохшееся тело. Такие, по выражению Догеля, «мумифицированные» жгутиконосцы всегда есть в прямой кишке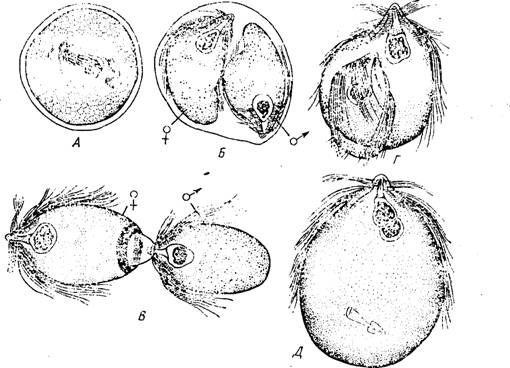
Рис. 33. Стадии полового процесса у жгутиконосца Trichonympha (по Кливленду). А — циста (деление ядра инцистированной особи); Б — сформированные гаметы, заключенные в оболочке цисты; В — втягивание мужской гаметы в цитоплазму женской гаметы; Г—Д — слияние мужского и женского ядер
термитов. По-видимому, именно они и служат источником заражения новых особей хозяина. Будучи заглочены, они без вреда для себя проходят через весь пищеварительный тракт термита и, попадая в прямую кишку, возвращаются там к активной жизни.
Размножение Hypermastigina осуществляется путем продольного деления, причем у них, как и у трихомонад, могут временно возникать формы с несколькими наборами органоидов. Половой процесс (рис. 33) известен только для тех видов жгутиконосцев, которые паразитируют в кишечнике древоядных тараканов. По Кливленду, половому процессу, который по времени связан с линькой хозяина, обязательно предшествует инцистирование. Органоиды пццнетпровавшейся особи разрушаются, а ядро делится карио- кинетическим путем. Вокруг ядер обособляется цитоплазм; формируются дпе гаметы — женская и мужская. Они остаю заключенными в цисте и внешне почти не отличимы. Лишь' ПО' эксцистирования происходит морфологическая дифференцироі гамет. В цитоплазме женской гаметы скапливаются интенеш окрашивающиеся гранулы, которые образуют в последней Yp тела широкое кольцо. Позади пего находится подвижный и клеіп цитоплазматический бугорок. Мужская гамета, лишенная э' образований, прилипает к клейкому бугорку женской гаметы и рез некоторое время втягивается внутрь ее цитоплазмы. Вслед тем происходит разрушение всех органоидов мужской гаметі слияние мужского и женского ядер. Синкарион претерпевает ме тическое деление.
Еще по теме ПОДОТРЯДЫ OXYMONADINA (HONIGBERG, 1964) И HYPERMASTIGINA (GRASSI ЕТ FOA, 1911):
- ПОДОТРЯД TRICHO.MONAD1NA (HONIGBERG, 1964)
- ПОДОТРЯД EIMERIIDEA. LEGER, 1911
- Г. С. ШИБРЯЕВА. СУДЕБНЫЙ ПРОЦЕСС ПО ДЕЛУ ВЕРХОВНОГО ГЛАВНОКОМАНДОВАНИЯ ГИТЛЕРОВСКОГО ВЕРМАХТА(Темплан 1964 г. Изд-ва ИЛ, пор. № 210), 1964
- ПОДОТРЯД HAEMOSPORIDIIDEA DAN1LEVSKIJ, 1885
- ПОДОТРЯД NEMATOCERA — ДЛИННОУСЫЕ
- ПОДОТРЯД TRYPANOSOMATINA KENT, 1880
- ПОДОТРЯД BOOONINA I10I.LANDE, 1952
- ЗНАЧЕНИЕ РЕВОЛЮЦИИ 1905–1911 гг.
- Тема 17. СССР в 1964-1991 гг.
- Революция в Иране 1905-1911