Происхождение и эволюция миксоспоридий
Миксоспоридии, судя по характеру вегетативных стадий, связаны в своем происхождении с классом саркодовых и, по-видимому, ведут начало от каких-то паразитических амеб[8]. Древние слизистые споровики, еще близкие к амебам, вероятно, имели вид мелких, подвижных плазмодиев, в которых формировалось небольшое количество просто устроенных спор.
Такие формы до наших дней не сохранились, но среди миксоспоридий, паразитирующих в желчном пузыре рыб, встречаются виды, характеризующиеся рядом очень примитивных признаков. Таковы представители рода Сега- іотуха, мелкие плазмодии которых содержат всего три-четыре ядра. Для другого рода — Sphaeromyxa — характерно примитивное строение спор (рис. 101), которые одеты грубыми толстыми створками. В их капсулах, расположенных на противоположных полюсах споры, заключены свернутые в рыхлую спираль толстые и короткие полярные нити.Миксоспоридии, паразитирующие в тканях, имеют, по-видимому, вторичное происхождение. На это указывают, прежде всего, гигантские размеры их плазмодиев, производящих миллионы спор (результат «отбора на плодовитость», столь характерного для па-разитов). Да и само строение спор, которые благодаря совершенной форме успешно противостоят давлению окружающих пошей (с. 174), а попадая в кишечник хозяина, надежно закрепляются там с помощью длинных п тонких стрекательных нитей, пжормт о проделанной ими эволюции.
Судя но тому, что морфологически наиболее примитивные мпк- соспоридии локализуются в желчном пузыре рыб, можно думать, что древние миксоспоридин паразитировали именно в этом органе. Высказанное предположение подтверждается н тем, что желчный
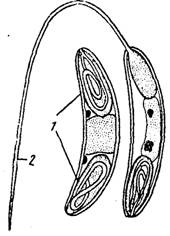
Рис. 101. Примитивное строение споры Sphaero- mijxa cottidarum (по Жукову, из кн. Шульмана): I — полярные капсулы. 2 — выброшенная стрекатель- ная нить
пузырь анатомически связан е двенадцатиперстной кишкой.
Благодаря этому, проглоченные рыбой паразиты могли легко попадать в желчный пузырь, перемещаясь по протокам. Освоенне же мпксо- спориднями других эндостаціні, расселение их по организму рыбы представляет собой результат длительной эволюции. Вероятно, первым шагом следует считать переход миксоспоридин к паразитированию в мочевом пузыре. В этом органе, морфологически сходным с желчным пузырем, паразиты нашли более или менее «привычные» условия существования. «Освоение» мочевого пузыря, а затем и связанных с ним протоков (мочеточников и мочевых канальцев) открыло для паразитов еще одни путь расселения — переход к паразитированию в мальпигиевых клубочках. Это и был, по существу, переход к тканевому паразитизму, облегченный особенностью самой ткани почек — рыхлой н хорошо снабжаемой кровью, а потому богатой питательными веществами. Следующим этапом эволюции миксоспоридин моіѵю быть поселение их на жабрах, органах с гораздо более плотными тканями, по физиологически близких к почкам (известно, что через жабры рыб удаляются мочевая кислота и аммиак, осуществляется активный транспорт ионов натрия).Дальнейшее расселение миксоспоридин, уже адаптированных таким путем к тканевому паразитизму, не представляет собой ничего неожиданного. Биологически более выгодная локализация в тканях, поставляющих паразитам неисчерпаемые пищевые ресурсы, естественно привела к вспышке видообразования. Не удивительно поэтому, что именно отр. Platysporea, к которому относится большинство тканевых видов; является едва ли не самым многочисленным отрядом миксоспоридин.
Описанный путь эволюции перехода миксоспоридин к тканевому паразитизму, по-внднмому, мог иметь место только у пресноводных рыб. С. С. Шульман связывает это с особенностями физиологии пресноводных н .мирских рыб. ІІЗПССТ- но, что все морские рыбы піпотоішчпы по отношению к внешней среде (морской цией воді,і в ночках хорошо развиты и очень многочисленны мнлышпіени клубочки. Такая морфологии органа по всей вероятности облегчила возможно, и.
перехода полостных миксоспоридин к паразитированию в своеобразной почечной ткани. Совсем иная картина характерна для морских рыб, у которых вода постоянно уходит из организма. В связи с этим у них преобладает не фильтрационная, а секреторная деятельность почек, направленная на сохранение воды. Почки морских рыб, выделяющие очень небольшое количество мочи, характеризуются малым числом, а подчас и полным отсутствием мальпигиевых клубочков. Органы такого строения навряд ли могли служить подходящим субстратом для поселения в них паразитов. И действительно, у морских рыб преобладают полостные виды миксоспоридин, тогда как тканевые представлены лишь небольшой по объему группой Multivalvulea. Огромное же большинство тканевых миксоспорилніі приурочено к пресноводным рыбам.иоде), тогда как пресноводные— напротив, гипертоничны. Это находит снос отражение и в строении их почек, органов, обеспечивающих не только удаление экскретов, по и водио-солспое равновесно (А. I. I ппецппскпіі). Ііочкн пресноводных рыб, в органним которых через кожные покровы непрерывно поступает вода, выделают большое количество пиюгонпчиои мочи. И еви.чи с активной фильтра
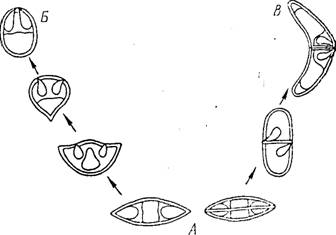
Рис. 102. Схема эволюции спор .миксоспоридин. Пе-- реход от спор типа Віроіагіа (.4) к спорам типа Platysporea (Б) и Eurysporea (В) (по Шульмаиу)
Паразитирование в тканях, оказывающих механическое давление на плазмодий и заключенные в нем споры, вызвало к жизни появление специальных адаптаций в*строении ' і і
спор.[Эти адаптации выражаются в приобретении линзообразной! форіиы п сводчатых створок, прочность которых усиливается пали-1 чием толстого шовного валика (с. 174). Эволюция строения спор і идет и в других направлениях. Это изменение формы створок и по- ■ явление выростов, обеспечивающих парение в толще йоды (с. 175),. а также тенденция к смещению полярных капсул. У более примитивных видов (отр. ІЗіроІагіа) полярные капсулы лежат па противоположных полюсах споры.
Однако необходимость усиления функции капсул, заключающейся в прикреплении споры к стейке кишечника хозяина, требует изменения в положении полярныхкапсул.
Если выстреленные полярные нити направлены в одну точку, они обеспечивают более надежное прикрепление споры, чем нити, направленные в противоположные стороны. Эволюционная тенденция к сближению полярных капсул в спорах миксоспорпдпп может быть проиллюстрирована морфологическим рядом (рис. 102), который позволяет приблизительно судить о направлении этого процесса в филогенезе. Сближение капсул могло осуществляться в плоскости шва (вдоль пего). Это должно было привести к появле- нию спор типа Myxobolus (отряд Platysporea). Другим путем могло быть смещение капсул в' плоскости, перпендикулярной шву, что могло также сопровождаться изгибанием створок и вытягиванием их в поперечном направлении, как у Ceratotnyxa (отр. Eurysporea). Наконец, полимеризация створок такого типа могла послужить основой для возникновения многостворчатых спор (подкл. Multival- vulea).
Таким образом, основными направлениями эволюции миксоспо- ридий следует считать тенденцию к увеличению продуктивности, способом осуществления которой явился переход к тканевому паразитизму, и тенденцию к совершенствованию строения спор, наи- лучшнм образом обеспечивающих заражение хозяина. ^
Еще по теме Происхождение и эволюция миксоспоридий:
- 6. «Новая теория о происхождении, разнообразии и эволюции живых организмов на Земле».
- Происхождение и эволюция паразитических Mastigophora
- 2. Первоначальное и производное происхождение гос-ва. Олигархическая теория происхождения государст-ва.
- 2. Эволюция ФЕНОТИПА и эволюция ГЕНОТИПА
- Помимо усилий по выявлению необходимых благоприятных условий для химической эволюции, создаются теории, в которых объясняются варианты химической эволюции при возможных неблагоприятных условия
- § 1. Основные теории происхождения государства Теологическая теория происхождения государства
- 2. Основные теории происхождения государства. Теологическая теория происхождения государства.
- § 2. Теории происхождения права Теологическая теория происхождения права
- 4. Прекращение правовой охраны наименования места происхождения товара и исключительного права на наименование места происхождения товара
- Вредные факторы производственной среды химического происхождения Источники вредных факторов химического происхождения на производстве
- Этика, доказанная в геометрическом порядке и разделенная на пять частей, в которых трактуется I. О боге II. О природе и происхождении души III. О происхождении и природе аффектов IV. О человеческом рабстве или о силах аффектов V. О могуществе разума или о человеческой свободе
- Эволюция психики.
- Процесс эволюции
- 11.11. Эволюция и революция
- 11.11. Эволюция и революция
- г) эволюция оценивания жизни
- Силы и механизмы эволюции
- К вопросу об эволюции современного человека To the question on evolution of the modern human
- "Хозяйство": историческая эволюция понятия
- "Хозяйство": историческая эволюция понятия