БИОХИМИЧЕСКАЯ СИСТЕМАТИКА
Отечественными биохимиками А.Н. Белозерским, А.С. Спириным, А.С. Антоновым и их последователями в начале 1970-х гг. XX в. были сформулированы представления об использовании генетических и биохимических показателей в оценке таксономического сходства живых организмов («Строение ДНК и положение организмов в системе», 1972), ныне получившие широкое распространение.
Со времен Эрнста Геккеля (1834-1919) известна триада сравнительно-филогенетических методов: морфологического, эмбриологического и палеонтологического. Исследователь, работающий классическими методами, сначала изучает морфологию и анатомию интересующих его объектов. Если черты строения органов, тканей и клеток не позволяют отнести их с достаточной уверенностью к тому или другому таксону, на помощь приходит эмбриология. В развитии организмов нередко сохраняются черты их отдаленных предков. Морфологические и эмбриологические данные там, где это возможно, дополняются палеонтологическими.
Комбинация этих методов — весьма мощный инструмент систематика-эволюциониста. Однако классические методы
42
УРОВНИ БИОРАЗНООБРАЗИЯ
пасуют именно в применении к низшим организмам. Морфологические признаки примитивных организмов за время их эволюции могли неузнаваемо измениться или, наоборот, дать многочисленные примеры конвергенции, а также параллельного развития. Кроме того, примитивные организмы именно в силу их примитивности, мало пригодны для изучения морфологии. Формы большинства бактерий (палочка, шарик, спираль) могли возникнуть самостоятельно в разных группах и многократно перейти друг в друга независимо на разных путях эволюции. То же относится к эмбриологическому методу: можем ли мы назвать «онтогенезом» деление бактериальной клетки или самосборку вирусной частицы? Палеонтологический метод также неприменим там, где речь идет об одноклеточных бесскелетных организмах: они практически не сохраняются в окаменевшем состоянии.
Из этого следует, что для решения проблем систематики низших организмов необходимы принципиально новые методы, позволяющие оценить степень родства, исходя из каких-то других признаков. Такими методами могут быть биохимические.Для понимания степени филогенетического родства организмов неоднократно использовались данные о составе и свойствах их алкалоидов, жиров, белков, путях синтеза тех или иных метаболитов. Широко известны также попытки применения иммунобиологических и иммунохимических методов, а также анализа элементного состава (Лебедева, 1999).
Однако подобные исследования имели весьма ограниченный успех и не получили признания у «классических» систематиков. Дело здесь не в консерватизме систематиков. Применявшиеся биохимические показатели были так же вторичны, как и морфологические. В конечном счете нет разницы между, допустим, числом щетинок у рачка и числом изози- мов его лактатдегидрогеназы — обе величины суть признаки фенотипа, а не генотипа.
По мнению А.Н. Белозерского и его учеников, переворот в биохимической систематике произошел после того, как американский ученый Э. Чаргафф в самом начале 1950-х гг. совершил открытие кардинальной важности, установив, что нуклеиновые кислоты, так же как и белки, видоспецифичны. На протяжении последующих 20 лет были получены данные, неопровержимо доказывающие, что нуклеиновые кислоты — не только материальная форма хранения наследственной ин-
43
ГЛАВА I
формации, но и те соединения, через которые осуществляется реализация наследственности в процессах развития организма. Крайне перспективно изучение нуклеиновых кислот с точки зрения познания основ эволюционного процесса — наследственности и изменчивости.
В первую очередь речь должна идти о дезоксирибонуклеиновой кислоте — ДНК, так как именно это соединение является материальным субстратом наследственности подавляющего большинства организмов (за исключением РНК-содер- жащих вирусов). В течение более 15 лет была проведена огромная работа по изучению первичной структуры ДНК различных бактерий, актиномицетов, грибов, водорослей, высших растений, а также позвоночных и беспозвоночных животных.
Полученный материал позволил сделать ряд важных выводов. Первый вывод состоит в том, что ДНК всего органического мира едина: во всех исследованных группах животных и растений она слагается из одних и тех же дезоксирибо- нуклеотидов, включающих два пуриновых основания (аденин и гуанин) и два пиримидиновых (цитозин и тимин). Для всего органического мира (за исключением вторичного явления — одноцепочечных ДНК некоторых вирусов) строго соблюдаются закономерности, подмеченные Чаргаффом («правила Чар- гаффа»), которые сводятся к следующему:1. Сумма пуриновых нуклеотидов равна сумме пиримидиновых нуклеотидов:
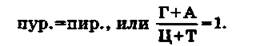
2. Содержание аденина равно содержанию тимина:
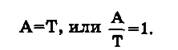
3. Содержание гуанина равно содержанию цитозина:
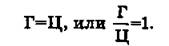
4. Содержание 6-аминогрупп равно содержанию 6-кето- групп:
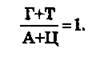
44
УРОВНИ БИОРАЗНООБРАЗИЯ
Г»
5. Содержание Г+Ц и А+Т может варьировать в довольно значительных пределах.
Поэтому для ДНК наряду с единым планом строения, обусловленным ее жесткой двуспиральной структурой, присуща возможность бесконечного варьирования состава и последовательности оснований.
Несомненно, возникновение ДНК в процессе происхождения жизни на Земле было важнейшим фактором дифференциации и становления новых видов. В структуре ДНК наследственная информация не только сохраняется наилучшим образом, но и может передаваться путем репликации от поколения к поколению. В бесконечной цепи следующих друг за другом поколений естественным отбором были выбраны структуры ДНК, обеспечивавшие не только стабильность наследственной информации (консерватизм наследственности), но и возможность вариации первичной структуры ее макромолекулы.
Из пятого правила Чаргаффа вытекает второй, не менее важный вывод, сделанный одновременно и независимо друг от друга А.Н. Белозерским и А.С. Спириным и французскими исследователями Ли, Валь и Барбю, а именно: процент пар гуанин—цитозин, изменяющийся в процессе эволюции, видоспецифичен и может иметь таксономическое значение. Впервые это было показано на ДНК бактерий, где мы имеем очень большие вариации нуклеотидного состава. Показатель
у бактерий может меняться от 2,57 (сарцина) до 0,45 (возбудитель газовой гангрены). Если этот показатель равен 1 (Г+Ц=А+Т=50%), такую ДНК называют эквимолярной. В ДНК ГЦ-типа гуанина и цитозина более 50%, для ДНК АТ- типа характерно обратное соотношение оснований.
Впоследствии вариабельность нуклеотидного состава ДНК была изучена у многих групп водорослей, грибов, вирусов, а также высших растений и животных. Анализируя эти данные, А.С. Антонов пришел к выводу о том, что вариабельность соответствует эволюционному возрасту таксона. Древние группы (вирусы, бактерии, грибы, водоросли) очень варьируют по составу ДНК. То же отмечается и у животных. Так, например, у древнейшей группы простейших содержание Г+Ц может меняться почти на 39%, у губок эта величина уже 25%, у хордовых— менее 6%.
45
ГЛАВА I
Не исключена возможность, что значительная вариабельность нуклеотидного состава ДНК микроорганизмов свидетельствует об их полифилии; в то же время она может быть лишь следствием длительности мутационного процесса. Любопытно, что ДНК практически всех высших растений и животных АТ-типа, причем нуклеотидный состав довольно стабилен. Специфичность в данном случае реализуется по линии нуклеотидной последовательности. Это обстоятельство ограничивает применение такого показателя, как нуклеотидный состав ДНК, у высших организмов. Тем не менее, использование данных о составе ДНК позволяет сделать более обоснованный выбор из числа имеющихся альтернативных схем макрофилогенеза.
А.С. Антоновым и Б.М. Медниковым (1972) методами вариационной статистики был проанализирован материал о составе ДНК около 2000 видов различных организмов. Из полученных данных сделан ряд любопытных выводов. Прежде всего можно заключить, что выделенные ранее формальные категории ДНК — ГЦ- и АТ-типа — не субъективны, а отражают реально существующую в природе закономерность: во всех наиболее изученных группах чаще всего встречаются ДНК, содержащие от 35 до 45 или от 50 до 60 мол.% ГЦ-пар нуклеотидов. Иными словами, мы имеем два модальных класса. Первый модальный класс (АТ-тип ДНК) прослеживается в группах хордовых и беспозвоночных животных, высших растений, грибов, сине-зеленых водорослей, бактерий и вирусов. ДНК второго модального класса (ГЦ-тип ДНК) отмечена у грибов, актиномицетов, водорослей, бактерий и вирусов. Самое удивительное то, что эквимолярный состав ДНК встречается относительно редко. Этот факт загадочен, так как ДНК с равным количеством всех четырех оснований должна обладать максимально возможной информационной емкостью (два бита на нуклеотид, согласно уравнению Шеннона).
Биохимики разработали методы сравнения организмов по набору аминокислот, а также по прямому сопоставлению молекул ДНК.
Второй подход сравнения последовательностей в ДНК более перспективен. Это так называемый метод молекулярной гибридизации. За последние годы он получил широкую известность среди микробиологов. Можно утверждать, что пер-
46
УРОВНИ БИОРАЗНООБРАЗИЯ
вые успешные попытки создать филогенетическую систему вирусов и бактерий основаны главным образом на применении этого метода.
Метод молекулярной гибридизации был разработан в США Хойером, Мак-Карти и Болтоном. Принцип метода сравнительно несложен. Как известно, нативная ДНК состоит из двух комплементарных цепей, соединенных водородными связями между аденином и тимином, и гуанином, и цитозином. В нормальных условиях клетки эта структура весьма устойчива. Если же повысить температуру, особенно в условиях низкой ионной силы, водородные связи между комплементарными цепями рвутся: ДНК денатурирует.
При медленном охлаждении начинается процесс ренатурации, воссоединения комплементарных цепей. Ренатурация приближается к стопроцентной только у гомогенных ДНК фагов. Уже у бактерий геном настолько неоднороден, что даже за большой промежуток времени не все последовательности могут найти себе комплементарные половины.Если денатурированную ДНК иммобилизировать осаждением на целлюлозный фильтр или включением в агаровый либо полиакриламидный гель, ренатурации не произойдет. Работая одним из основных методов, процесс гибридизации проводят в колонке с гелем, в который «вплавлена» денатурированная ДНК. Затем через колонку пропускают раствор ДНК, также денатурированной, но, кроме того, подвергнутой деградации (молекулы ДНК легко разрываются на части; этот процесс называют деградацией и обычно в качестве деградирующего агента применяют ультразвук). Короткие фрагменты деградированной ДНК легко проходят через поры геля и могут соединяться с иммобилизированной ДНК, образуя реассоциированные двойные цепи. Низкомолекулярную ДНК обычно метят радиоактивным фосфором, углеродом или тритием. В дальнейшем надо лишь вымыть из колонки неприсоединившуюся меченую ДНК. Весьма полезен прием, заключающийся в обработке препарата специфической ДНК-азой, расщепляющей одноцепочечную ДНК, но не трогающей двухцепочечную. В результате в геле остаются только гибридные цепи (в тех случаях, когда ДНК иммобилизированная и меченая происходят из разных источников). О проценте гибридизации мы можем судить по активности конечного продукта.
47
ГЛАВА I
Болтон и Мак-Карти прекрасно отдавали себе отчет в том, насколько разработанный метод может быть цепным именно для систематики. В конечном счете это прямое определение близости геномов друг к другу. По ряду причин методу молекулярной гибридизации «нестрашны» конвергенция и параллельное развитие — проклятие всех ныне существующих методов определения сходства организмов. Например, вполне вероятно конвергентное развитие макроструктур у неродственных форм. Весьма часто и конвергентное возникновение сходных по функции белков: лизоцимы фагов и высших организмов, несомненно, не родственны. Однако конвергентное возникновение идентичных аминокислотных последовательностей уже крайне маловероятно. Еще менее вероятно подобное возникновение идентичных участков цепей ДНК, даже если они кодируют сходные по составу белки. Ведь генетический код вырожден и большинству аминокислот может соответствовать несколько триплетов. Поэтому молекулярная гибридизация — весьма многообещающий метод для выявления полифилетических таксонов.
Даже в столь хорошо изученной группировке, как млекопитающие, этим методом подтверждена полифилия отряда грызунов, многими систематиками давно разделяемого на отряды зайцеобразных и двурезцовых. Известно, что дискутируется вопрос о разделении на самостоятельные отряды зубатых и беззубых китов, а также ластоногих и насекомоядных.
Однако возникли не только технические, но и методические трудности. Уже первые работы по гибридизации ДНК показали, что у высших организмов геном неоднороден. В самом первом приближении его можно разделить на две фракции: фракция уникальных последовательностей и фракция повторяющихся последовательностей. Не менее интересны сателлитные-фракции ДНК — оказалось, что концентрации их варьируют очень сильно даже у близкородственных видов.
Комбинируя условия выделения, фракционирования и гибридизации ДНК, в принципе можно сопоставлять геномы в пределах таксонов любого ранга, вплоть до вида и в конечном счете весьма облегчить работу систематика, предоставив ему уникальные данные биохимического анализа. Методика молекулярной гибридизации, хотя и не представляет собой
48
УРОВНИ БИОРАЗНООБРАЗИЯ
ничего сложного, чрезмерно трудоемка. Обнадеживает то, что она поддается автоматизации, а в ряде лабораторий США и Западной Европы поставлена «на поток», стала рутинной процедурой, хотя и довольно дорогостоящей.
Впрочем, уже достаточно давно метод молекулярной гибридизации в сочетании с определением нуклеотидного состава и анализом изоплит дает результаты и не только при решении частных проблем систематики. Так, группой А.С. Антонова были проведены опыты по изучению степени гомологичности первичных структур геномов у однодольных растений и разных классов хордовых животных (главным образом птиц, рептилий). Следует остановиться на одном выводе из серии этих работ, имеющем для построения систем принципиальное значение.
Речь идет о разномасштабности таксонов в пределах различных групп растительного и животного мира. Известно, что все системы построены по иерархическому принципу: виды объединяются в роды, роды в семейства, семейства в отряды (или порядки в фитосистематике) и т. д., вплоть до царств живой природы. Возникает вопрос: соответствует ли род, например, однодольных растений роду птиц или млекопитающих? Ответить на этот вопрос трудно, так как если границы таксона систематики обычно очерчивают правильно, то ранг присваивается довольно произвольно и имеет тенденцию повышаться в связи с изученностью группы. Ясно, однако, что если таксоны равномасштабны, то и степень гомологичности первичных структур ДНК у образующих их видов должна быть одинаковой.
Однако первые опыты по гибридизации ДНК показали другое — разномасштабность таксонов в системах разных групп растительного и животного мира. Так, например, меченая ДНК домашней курицы имеет приблизительно 25—38% гомологичных последовательностей с ДНК птиц других отрядов (исследовались воробьиные, трубконосые, попугаи, голуби и дневные хищники). Приблизительно Такая же межотрядная степень гомологии ДНК установлена Мак-Карти и Болтоном для класса млекопитающих. Степень гомологичности последовательностей в ДНК представителей разных классов позвоночных, естественно, ниже (у ДНК курицы и варана, например, 10,5%). Эти участки генома, по всей вероятности, кодируют наиболее общие признаки, присущие всем хордовым.
49
ГЛАВА I
Совсем другие цифры получены для однодольных растений. Виды, относимые к разным семействам порядка лилиецветных, имеют лишь до 10% гомологичных структур ДНК. Иными словами, семейство в группе однодольных растений по рангу должно быть приравнено к классу хордовых животных! Представители разных порядков однодольных обладают еще более гетерологичной ДНК, а между ДНК однодольных и двудольных растений иногда вообще не образуется гибридных молекул. О субъективности ранга систематических категорий говорилось уже неоднократно, но теперь мы можем количественно оценить, насколько велика эта субъективность. Во всяком случае эти данные свидетельствуют о различиях кардинального свойства в путях и темпах эволюции геномов.
Еще по теме БИОХИМИЧЕСКАЯ СИСТЕМАТИКА:
- 3. Систематика международного права
- Поиски систематики человека
- 3. Практики-систематики
- Систематика любви.
- Систематика грибов
- Систематика дрожжевых грибов
- § 61. Методологическое значение систематики феноменологических редукций
- От гена к психике. Структурные, биохимические и физиологические влияния.
- 13.5. Раскрытие роли гипогликемии как одного из биохимических стимуляторов агрессивности
- 3.1. Биохимические показатели сыворотки крови у испытуемых разных типов телосложения.
- 7.4 Влияние лактулозы на биохимические свойства микрофлоры и процессы структурообразования в кисломолочных продуктах
- 1.3. Влияние физической нагрузки на биохимические показатели крови у людей разных соматотипов.
- Морфологические и биохимические критерии гипоксии при разработке материалов для МКЛ
- 15. Задержанное развитие как вариант психического дизонтогенеза. Систематика ЗПР.
- I. ВВЕДЕНИЕ Структура учебного курса «Гражданское право (часть 2)» основывается прежде всего на систематике второй части