ГОМІНОЇДИ
Гоміноїди з’явилися в Африці, очевидно, в кінці олігоцену; їхніми предками були маленькі антропоїди сімейства пропліопітеків. Приблизно 30-25 млн. р. тому проходить поділ вузьконосих мавп на дві гілки - церкопітекоїдів (Cercopithecoidea) чи собакоподібних (сучасні павіани та ін.) і гоміноїдів (Hominoidea) чи людиноподібних (сучасні шимпанзе та ін.).
Існують припущення, що такий поділ викликаний переходом собакоподібних мавп на харчування переважно листям дерев і кущів, в той час як людиноподібні мавпи, крім листя, харчувалися також плодами дерев, насінням рослин і горіхами. Такий харчовий раціон з часом відбився на будові кутніх зубів гоміноїдів - т. зв. “візерунок дріопітека” (чотири жувальних виступи на молярах церкопітекоїдів) є в багатьох випадках головною ознакою віднесення решток до тих чи інших родів і сімейств (зуби вкриті твердою емаллю і є найчастішими знахідками скелетів гоміноїдів). Другою характерною їхньою ознакою є, як уже зазначалося, відсутність хвоста, короткий і широкий тулуб по відношенню до кінцівок і специфічна будова плечового суглобу. Останнє давало можливість сидіти на гілках дерев і пересуватися способом брахіації - розгойдування і перестрибування з гілки на гілку.Сьогодні в Африці, Азії і Європі нараховують більше ЗО родів різних міоценових гоміноїдів, які зайшли у своєрідний еволюційний тупик, не давши нащадків. Однією з найраніших форм вважають рід камояпітека(Kam oj ар і th ecus) з пізньоміоценового місцезнаходження Лосі- док в Кенії, що має чітке датування в межах 27-24 млн. р. тому. Ще одне місцезнаходження в Кенії - Меса Бридж має хронологічну дату 20 млн. р. [Вишняцкий, 2005, с. 34].
Пропліопітек, парапітек,(Propliopithecus, Parapithecus), можливо, входять уже до складу вузьконосих. Найдавнішим з них є олігопітек - Oligopithecus (вагою всього 0,6-1,0 кг), трохи пізніше з’являється крупніший “плодоїдний" тип єгиптопітєка - Aegyptopithecus (6 кг вагою), який належить уже до гоміноїдної гілки еволюційного розвитку.
Своєрідною скарбницею ранніх гоміноїдів є місцезнаходження Фаюм у Єгипті - пустельна місцевість за 100 км від Каїру. У відкладах пізнього еоцену і олігоцену (50-25 млн. р. тому) у Фаюмі вчений ПІльосер ще у 1910 р. відкрив і описав багато видів, розподіливши їх на дві великі групи: парапітеків та пропліопітеків, назвавши їх надсімействами. Парапітеціди включають: Parapithecus, Apidium (апідіум) і Qutrania (кватранія) з Африки, Amphipithecus (амфіпіте- кус) з Бірми. Вузьконосі (Catarrhini) мавпи вищого ґатунку могли походити від цих примітивних істот. Це викопні предки мавп, що жили в олігоцені - 35-30 млн. р. тому. Мали малі і середні розміри, величиною з мавпу ревуна, мали видовжену, опущену вниз морду. Знайдено зуби,

Рис. 9. Карта Старого Світу з окресленою зоною проживання людиноподібних мавп, головних знахідок гоміноїдів і гомінідів (австралопітеків)

Рис 10 Поширення гоміноїдів на карті Старого Світу
фрагменти черепа, кілька фрагментів кінцівок, які мало що можуть сказати про спосіб локомоції (пересування) цих істот. Джон Нейпір схильний вважати, що вони пересувалися на чотирьох кінцівках і були брахіаторами. Невідомо, чи ці істоти взагалі були предками людини і людиноподібних мавп.
В Африці в період раннього міоцену проживали дімнопітеки(Limπopithecus) і мікро- пітеки(Mikropithecus). Важливе значення має знахідка моропітека (Moropithecus) з Уганди (20,6 млн. р. тому). Це були великі плодоїдні істоти вагою 40-50 кг. їхній скелет був уже вертикальний і вони могли, розгойдуючись, стрибати з гілки на гілку. Крім того, з’являються різні інші види - дендропітеки (Dendropithecus macmesi) в Африці вагою 9 кг (22 млн. р. тому) чи плюпіте- ки (Pliopithecus vindobonensis) з Девінска Нова Весь поблизу Братислави.
Вищі примати з’являються в міоцені.
В цей час клімат змінюється до холоднішого, стає сухішим, ліси поступово змінюються на лісостеп [5machel, 2005, р. 6]. В ранньому міоцені пропліопітеки трансформуються у рід гібонів. Найдавніший типовий гібон походить від виду діони- сопітека(Dionysopithecus) з Китаю (12 млн. р. тому), а найпізніший від виду крішнапітека (Krishπapithecus) з Індії (8 млн. р. тому); раніше вони були описані як пліопітеки (Pliotithecus).Дендроттек(Dendropithecus) жив у ранньому міоцені (25-20 млн. р. тому). Останки знайдені на території сучасної Кенії. Розмірами був не більший від гібона, мав тонку структуру, як у павукоподібної мавпи (коати). Лазив, стрибав і висів на деревах.
Пліопітек(Pliopithecus) жив у міоцені (20-15 млн. р. тому). Одним з першим відкритих у Європі видів антропоїдів був Pliopithecus

Рис. 11 Схема еволюційної історії приматів
antiquis, описаний Ларте ще у 1838 р. у Франції. Раніше вважався предком гібона, мав такі самі розміри і подібну мордочку - великі очі і великі гострі ікла, але на відміну від гібона він не мав таких великих кінцівок і, можливо, мав хвіст.

Рис. 12. Реконструйований череп проконсула
Проконсул(Proconsul). Одним з найбагат- ших сімейств періоду пізнього міоцену в Африці та Євразії були Proconsulidae, що датуються ча* сом переважно 15-5 млн. р. тому, хоч є дані і про 22 млн. р. тому. “Проконсул” - назва походить від клички шимпанзе “Консул” з лондонського зоопарку. Префікс “про-” означає “до ” - тобто, це істоти, що існували в період “до шимпанзе”, були давнішими. Важливі знахідки решток проконсулів знайдені Мері Лікі в 1959 р. в Кенії, та Аланом Уокером і Мартіном Пікфордом у 1980 р. в Найробі. Об’єм мозку у нього порівняно великий, у морфології скелету простежуються риси звіроподібних і людиноподібних мавп.
Черепна коробка займає проміжний розмір поміж черепами павіанів і шимпанзе. В конфігурації лицевого скелету переважають церкопітекоїдні особливості [Бунак, 1959). Кістки кінцівок і стопи мають прогнатний вигляд. Деталі будови черепа і зубів вчені добре вивчили на знахідках з побережжя оз. Вікторія в Африці. Ранньоміоценова вулканічна активність сприяла захоронению і збереженню цих решток. Лава є ідеальним консервацій ним матеріалом для викопних кісток як тварин, так і людини. Враховуючи величину зубів, наймасивніший вид - проконсул майор сягав розмірів горили і фактично міг бути його предком, інший вид - проконсул африканський міг бути предком шимпанзе. Не можна виключати, що останній вид започаткував гоміноїдну лінію людини. Більшість спеціалістів допускають, що це сімейство могло бути спільним предком для розвитку усіх подальших гоміноїдів.Дріопітикові.(Dryopitheciπae, Dryopithe- CUδ) жили в середині міоцену (13-8 млн. р. тому). Рештки знайдені в Європі, зокрема в Франції. Це також одна з перших відкритих викопних форм мавп (1856 р.). Дріопітеками (лісовими мавпами) їх було названо тому, що перші знахідки трапились у геологічних шарах, які вказували на дуже теплий клімат і тропічне лісове середовище. В наш час знахідки відомі в Європі, Африці, на Близькому Сході, в Китаї, Росії. У

Рис. 13. Гоміноїди міоценового періоду
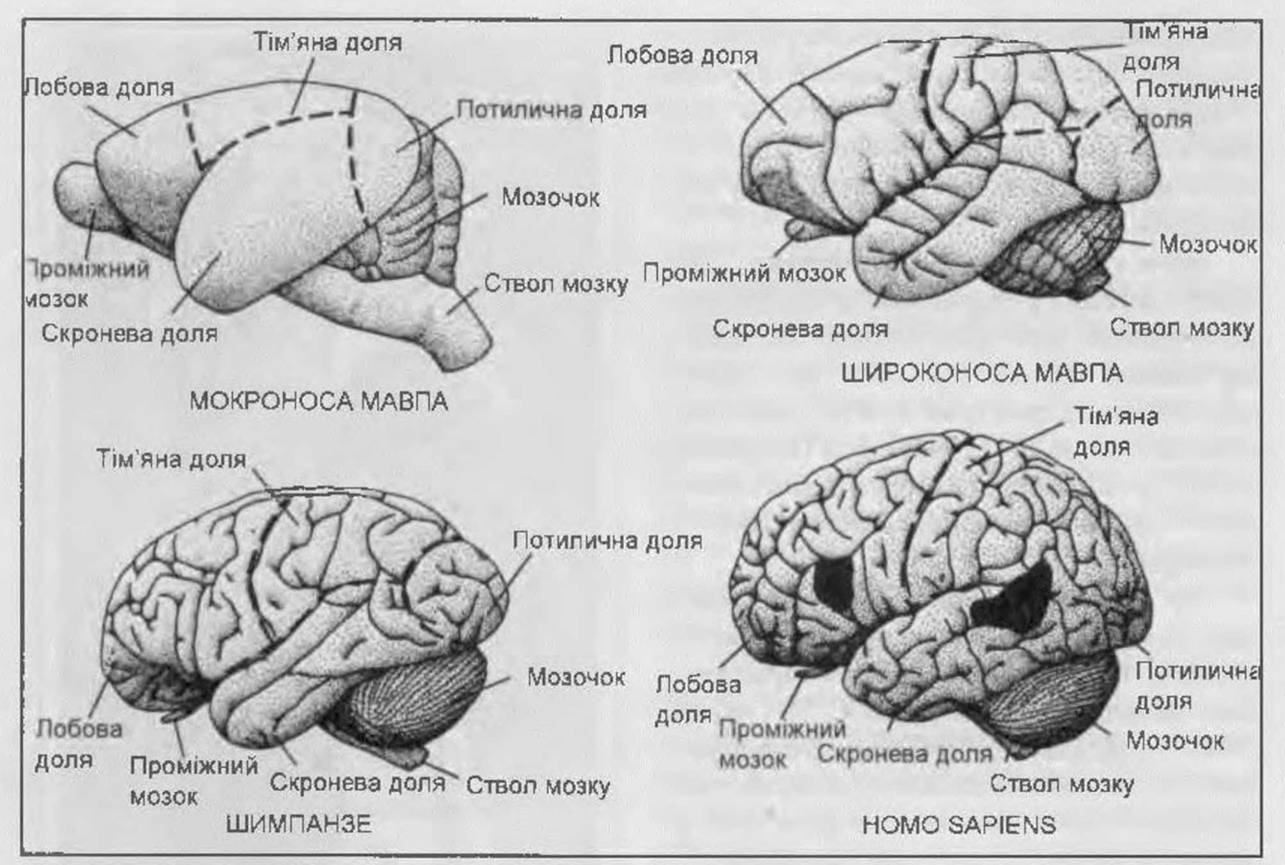
Рис. 14. Порівняльна характеристика мозку різних приматів і людини
1996 р. в Іспанії трапилися знахідки добре збережених частин скелету Driopithecus laietanus на місцезнаходженні Сан Лобатерес.
Це була потужна група істот, що заселяла зону теплих і помірних лісів, не доходячи до тропіків. Дріопітеки відзначалися найбільше випрямленою поставою тіла серед усіх інших гоміноїдів того часу. їхні плечі були порівняно широкими, що давало можливість робити різні вправні рухи руками, кисть була широка і пристосована до захвату різних предметів.
Можливо, були предками однієї з гілок сучасних людиноподібних мавп.У цих істот руки, плечовий пояс і зап’ястя пристосовані до того, щоб розгойдуватися на деревах; грудна клітка широка, поясниця вкорочена, але основа спини видовжена, розширені тазові кістки, г копчик. Зуби і череп із широкою і часто видовженою лицевою частиною дуже подібні до зубів інших гоміноїдів Старого Світу. Розміри тіла і відносна величина мозку були більшими.

Рис. 15. Черепи різних гоміноїдів міоцену
Вони з’явилися в Африці, очевидно, в міоцені і прийшли у Європу під час пересихання доісторичного моря Тетіс. Лазили по деревах, харчувалися плодами, оскільки їхні корінні зуби з тонким шаром емалі не були пристосовані до пережовування грубих харчів (наприклад, зерен). Дріопітек разом з африканським кеніапі- теком є одним з ранніх представників сімейства Hominidae.
За двома винятками, дріопітеки відомі лише за знахідками фрагментів зубів та щелеп, можливо тому їхня морфологічна характеристика неодноразово змінювалася. Е. Саймонс (1965) та Дж. Сімлсон (1967) визначали дріопітеків як особливе підсімейство сімейства пон- гід (вищих мавп) і відносили до дріопітеків усі міоценові форми, за винятком ореопітеків. За схемою Сімпсона, дріопітек фонтанов - вихідна форма горил, сівапітек - орангів, пліопітек - гібонів.
Рамапітеки(Ramapithecus) найпоширенішими були на території сучасного Пакистану і Непалу в період 13-8 (15-7) млн. р. тому. Вперше виявлені на Сиваликських пагорбах в Індії. Представлені знахідками лише щелеп із зубами. Іноді їх називають ще сугрівапітеками і брахма- пітеками.
Цікавою е історія відкриття цього гоміної- да. Ще в 1878 р. геолог В. Теобальд зазначив, що в цьому регіоні знайдено викопні рештки людиноподібних мавп. Подальші пошуки підтвердили слушність його інформації. Тут відкрито цілу колекцію різних кісток мавп неогенового періоду.
Серед них і рамапітеки (дослівно - мавпа Рами). Рама - герой епосу “Рамаяма", який за допомогою мавпячого війська переміг армію рак- шасів - злих демонів.Вид вперше був описаний Г. Дьюісом у 1934 р. і викликав дискусію серед спеціалістів. За його описом, вони мали “малі зграбні щоки і зуби, нижня щелепа коротка, зуби наближені до людських, порівняно невеликі різці, ікла дуже
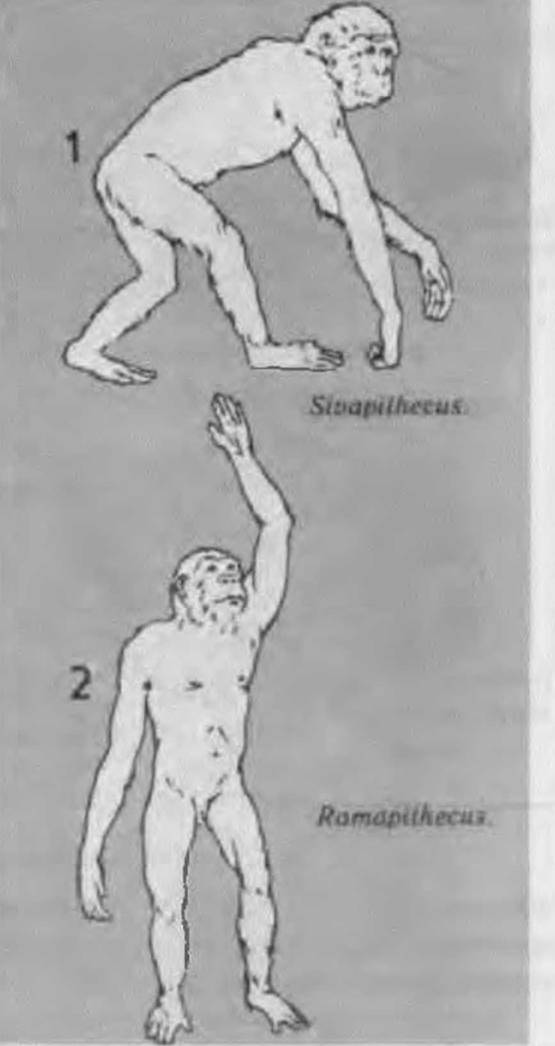

Рис. 16. Умовний вигляд сіва- та рамапітеюв

Рис 17. Череп сівапітека

Рис. 18. Способи пересування (локомоції) різних тварин і людини
малі”. Рамапітек жив власне у той час, коли стався той вирішальний крок, який розділив лінію людини і сучасних людиноподібних мавп. Поряд з тим, рамапітек має більше мавпячих рис, ніж людських. Жили вони посеред рідколісся і частину дня проводили в кронах дерев. “Однак на відміну від шимпанзе, - писав відомий палеоантрополог К. Хауел, - які харчуються переважно м’якими овочами, рамапітеки перейшли уже до твердих харчів - зерен злаків, коріння, насіння, горіхів. На це вказує товстий шар емалі на зубах і сліди тертя на кутніх зубах”. Оскільки рамапітек частіше використовував кутні корінні зуби, то прогнатизм його морди (лиця) зменшився. Спочатку його так і назвали - Ramapithecus brevirostris (короткопиский) [Kondratow, 1983, s. 76-781.
Внаслідок порівняльного аналізу Е. Саймонс [Simons, 1968} виділив особливий рід рамапітеків з типовою формою - рамапітек пенджабський. Зубна дуга у рамапітека у формі латинської літери “V”, у людиноподібних мавп - у формі літери “U”, у людини - у формі параболи [Ламберт, 1991, с. 83|.
Рамапітек, можливо, є зменшеним варіантом сівапітека. У нього була вкорочена лицева частина і дрібніші плоскіші зуби з порівняно великою поверхнею кусання, ніж у людини і у людиноподібних мавп. Виявлені глибокі щелепи, що були коротші і гостріші, ніж у людини і дріопітеків. Вага - 14 кг. Раніше рамапітеків вважали прямими предками гомінідів пізнішого часу. Від цих істот велася лінія людиноподібних мавп і людини.
Сіваттеки(Sιvapιthecus) - Пакистан, Індія, Непал, Туреччина. Жили в міоцені 15-9 (13- 5) млн. р. тому. Перші знахідки також трапилися на Сиваликських пагорбах в Індії. Це знахідки щелеп із зубами. Скелетні рештки рамапітеків, сівапітеків і дріопітеків в деяких випадках знайдені на одній і тій же території, що не виключає паралельного розвитку деяких міоценових груп. Треба врахувати, що у загальному ареалі види в період найвищої радіації могли знаходитися в різних екологічних нішах.
Сівапітек мордою був подібний до орангутангів, кістками стопи - до шимпанзе, міг маніпулювати зап’ястями. Міг лазити по деревах, висіти на гілках і ходити на чотирьох кінцівках. Був невеликий на зріст. Його крупні корінні зуби, вкриті товстим шаром емалі, свідчать, що ці тварини існували уже на відкритих місцевостях, харчувалися сухою і грубою їжею. Можливо, це був наслідок пристосування до умов відкритої лісової місцевості.
Сівапітека, як і рамапітека, ще недавно відносили до безпосередніх предків людини і лю- диноподібнихмавп.
Анкарапіпек(Ankarapithecus) знайдений у Туреччині. Датується міоценовим періодом - 9,8 млн. р. тому. Деталізовані дані опубліковані під кінець 1996 р. В анатомії черепа простежується суміш рис горили та орангутанга, (
Ореопітек(Oreopithecus) жив'ІМилйї р. тому, відкритий у 1872 р. в шарах верхнього мі-

Рис 19. Стадії еволюції антропоїдів та гоміноїдів (умовна послідовність)
оцену Північної Італії (декілька десятків кісток скелету багатьох особин). Розміри тіла відповідають розмірам середнього шимпанзе. Череп довгий і низький, з виступаючим надбрів’ям, лицевий скелет невисокий, слабо прогнатний. Кутні зуби з малою товщиною емалі, різці невеликі.
У будові кінцівок є подібність до церкопіте- кових і до гомінідів. Його зубна система змішана
- має риси первісні і похідні. Тулуб ореопітеків короткий, грудна клітка широка, руки довгі, а ноги короткі. Локомоція в значній мірі була двонога. Жили в умовах болотистих лісів. Його еволюційне місце не встановлене. Г. Швальбе виділив ореопітеків в особливе сімейство серед вузьконосих.
Сім пеон об єднує їх з нейтральними олігоценовими апідіумами. Б. Кемпбел [Campbell, 1961] і ІП. Штраус визнають ореопітеків особливим сімейством надсімейства Гомінідоідеа [Бунак, 1980].
У 2003 р. у Таїланді знайдено новий вид - люфенпітек (Lufengpithecus - 13 млн. р. тому), який за морфологією подібний до орангутанга (хоча сьогодні більшість дослідників виводять орангутанга від сівапітека). Отже, азійські сі- валітециди, включаючи люфенгпітека і анкара- пітека, прогресували в бік розвитку біологічної спеціалізації до орангів (?), а не до австралопітеків.
Африканські і європейські сіва/рама/піте- ціди розвивалися у двох напрямках локомоції
- на чотирьох кінцівках (quadrupedal), представниками якого стали шимпанзе та горила, і двоногого ходіння - оррорін, австралопітеки і людина.
Азійські сіва/рама/пітеціди також коево- люціонували у двох напрямках - на чотирьох кінцівках, як гігантопітеки, і в брахіаторному вигляді, як орангутанги [Smachel, 2005, р. 12].
Доказами спільного походження людини і людиноподібних сучасних мавп є подібність у будові тіла, поведінці і, перш за все, в біохімічних сполуках, з яких складається організм. Біо- молекулярні і генетичні дослідження вказують, що найближчими нашими родичами є людиноподібні мавпи Африки - найперше шимпанзе, потім горила. Один із дослідів показав, що ДНК людини і шимпанзе подібні на 98%. Крім цього, молекулярно-генетичні дані говорять про те, що еволюційні лінії людини і людиноподібних мавп розійшлися всього 7-5 млн. р. тому, значно ближче до нашого часу, ніж це вважалося раніше.
2.5.
Еще по теме ГОМІНОЇДИ:
- ЗМІСТ
- Педагогіка. Інтегрований курс теорії та історії: Навчально- методичний посібник: У 2 ч. / За ред. А.М. Бойко. — Ч. 2. — К.: ВІПОЛ; Полтава: АСМІ,2004. — 504 с., 2004
- Кармазин Ю.А., Стрельцов Е.Л. и др.. УГОЛОВНЫЙ КОДЕКС УКРАИНЫ. КОММЕНТАРИЙ. Харьков-Одиссей, 2001
- ПРЕДИСЛОВИЕ
- РЕДАКТОРСКАЯ СТАТЬЯ
- ОБЩАЯ ЧАСТЬ
- Раздел I
- ОБЩИЕ ПОЛОЖЕНИЯ
- Статья 1. Задачи Уголовного кодекса Украины
- Статья 2. Основание уголовной ответственности
- Раздел II ЗАКОН ОБ УГОЛОВНОЙ ОТВЕТСТВЕННОСТИ
- Статья 3. Законодательство Украины об уголовной ответственности
- Статья 4. Действие закона об уголовной ответственности во времени
- Статья 7. Действие закона об уголовной ответственности в отношении преступлений, совершенных гражданами Украины и лицами без гражданства за пределами Украины