Модель рынка бройлеров
Каждое утро и каждый вечер... кур... на столе
(13:11 2-я Паралипоменон) внутренности... печени... почки... кур...
(29:13 Исход) Не носите... кур... не могу терпеть
(1:13 Исаия) возьми...
кур... внутренности (29:22 Исход) И вывел всех... кур...(23:8 4-я Царств)
Как известно, любой живой организм в начале жизни набирает вес, затем скорость набора веса падает и, в дальнейшем, вес стабилизируется: сколько его ни корми, а, в среднем, выше биологической нормы веса набрать не удастся. Затраты кормов идут "впустую" (с т.з. набора веса). В этом "утилитарном" смысле на рынке бройлеров естественно возникает задача: как надо кормить птиц, сколько закупать цыплят, и в каком возрасте их отправлять в магазин, чтобы получить наибольшую прибыль. Сделаем очевидные допущения, что все купленные цыплята доживают до магазина, максимально возможный биологический вес одинаков у всех особей и, что вес цыплёнка по сравнению с максимальным весом особи мал. Для упрощения положим, что птицефабрика закупает ежедневно цыплят, кормит их, и через время: T продаёт в магазин, т.е. на фабрике в равномерном количестве имеются цыплята-куры всех возрастов в интервале времени их жизни {0...Т}. Пусть спрос равен предложению. Введём обозначения:
- m = m(t) - это зависимость массы особи m [кг] от времени t. Относительно характера этой зависимости отметим следующее. При t = О мы имеем массу цыплёнка т0, т.е. т(0) ξ т0. При t => 00 вес птицы (сколько ни корми) должен стабилизироваться, на некотором среднем её уровне, т°°, т.е. т(°°) ξ т°°. Ещё одно замечание. В начале роста цыплёнок набирает вес не так стремительно, как в среднем возрасте. Поэтому в качестве: m(t) можно предложить, например, такую функцию: m(t) = А/[В + Exp(-t/C)], где параметры А, В и C можно определить из следующих соображений. При: t = О имеем вес цыплёнка, т.е.
т0 = А/(В + 1). При: t => °° имеем явное соотношение: т°° = A/В. Значение: т°° можно определить экспериментально, для чего группу бройлеров кормим, например, в течение года, а после этого находим средний вес особи из этой группы. Определение параметра C (с размерностью времени) можно также найти экспериментально. Для этого надо кормить группу бройлеров и ежедневно определять их средний вес. Отмечаем время: Т1/2, когда средний вес достигнет половины максимального. После этого будет иметь место соотношение: 0.5*m°° = А/[В + Exp(-Ti/2/C)]. Решая совместно эти три уравнения, получим: В = m0/(m°° - m0); A = m°°*B; C = TWLn(IZB). Кстати, отмечу, что точка: TW является точкой перегиба для кривой m(t), т.е. в этой точке скорость роста массы птицы максимальна: ITi(TW)' = 0.25*А/(В*С). Например: ш0 = 0.1 кг; т°° = 3 кг; TW = 40 дней. Тогда: В « 0.0345, А « 0.1034 кг, C « 11.88 дня, и, соответственно наибольшая скорость набора веса будет: т(40)' = 0.25*0.1034/(0.0345* 11.88) « 0.0631 [кг/день].- К (или, при потерях поголовья, - K0) - количество покупаемых фабрикой цыплят [шт/день];
-S- себестоимость, или закупочная цена одного цыплёнка [руб/шт];
- Κ·Τ - полное количество птицы на фабрике, или ёмкость птицефабрики [шт];
- а - потребительная стоимость (прибыльность) потребления мяса бройлеров [руб/кг];
-X- отпускная цена мяса бройлеров с фабрики в магазины [руб/кг];
- μ - максимальный спрос на мясо бройлеров, при бесплатной его раздаче [кг/день];
-λ·Δί - вероятность потери (смерти) птицы на интервале времени: At [1/день].
А теперь составим баланс прибылей-убытков фабрики, как функцию параметров: х, К и Т, и попытаемся провести оптимизацию её работы в плане получения наибольшей прибыли.
Затраты на ежедневное приобретение фабрикой цыплят тривиальны, это: Qi = K*s.
Затраты на содержание фабрики, очевидно, они пропорциональны количеству бройлеров в ней: Q2 = α·Κ·Τ, где: а - некоторый коэффициент, который отражает удельные затраты на одно "птице-место" в день (персонал, освещение-отопление-канализация, амортизационные отчисления, охрана, ветеринария, арендные выплаты, проценты по кредитам и пр.).
Естественно, что при наличии птиц разных возрастов и разной массы, затраты кормов тоже различаются: цыплёнку в день надо меньше корма, чем взрослой особи. Примем в нулевом приближении, что расход корма пропорционален массе птицы. Поскольку масса у птиц всех возрастов различна, то общую биомассу птиц: Ms найдём интегрированием: Ms = K»J m»dt ξ K*A*(C/B)»Ln{[1 + В»Ехр(Т/С)]/(1 + В)}, где интегрирование по t проводим в пределах: {0...Т}. Откуда затраты на корма по всей фабрике будут: Q3 = β·Κ·Α·0/Β·Ι_η{[1 + В»Ехр(Т/С)]/(1 + В)]}, где: β - коэффициент, отражающий удельные (на 1 кг массы птицы) затраты на корм в день. Размерность у коэффициента: β равна [руб/(кг»день)], а для коэффициента: а - [руб/день].
Если принять зависимость спроса от цены экспоненциальной, то ежедневная выручка от продаж будет: Q4 = р*х*Ехр(-х/а). Следовательно, прибыль фабрики, как разность выручки и затрат составит: Q = Q4 - Qi - Q2 - Q3. К этому уравнению надо добавить уравнение связи параметров модели, отражающее тот факт, что количество покупаемых цыплят должно быть равным количеству продаваемых бройлеров (потери поголовья, напоминаю, отсутствуют). Это уравнение имеет вид: р»Ехр(-х/а) = К»т(Т), где левая часть уравнения это спрос [кг/день] на мясо птиц, а правая часть - это предложение [кг/день] птицы от птицефабрики. Исключив из выражения прибыли: Q = ... с помощью уравнения связи параметр К, получим выражение: Q(x, Т) = р*Ехр(-х/а)*[х - (s + α·Τ + β·1 m»dt)/m(T)]. Дифференцируя его по переменной: х, и приравнивая производную к нулю, получим уравнение, для определения оптимальной цены птицы, как функцию её "времени жизни": х(Т) = a + (s + α·Τ + β-J m*dt)/m(T). Подставив это значение: х(Т) = ... в уравнение для прибыли: Q = ... и дифференцируя уже по параметру T с
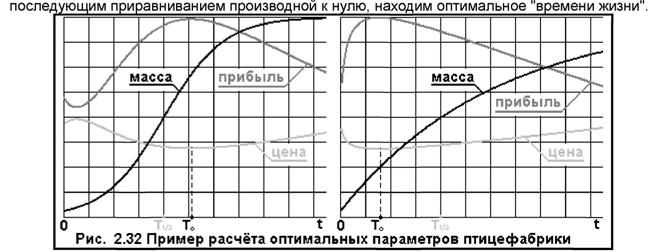
На Рис. 2.32 слева приведен графический способ поиска оптимальных параметров для птицефабрики.
Зная зависимость: m(t), и, задавая поочерёдно, значения: t, рассчитываем значение оптимальной цены: x(t) = ... и прибыли: Q = ... до тех пор, пока прибыль не начнёт снижаться (для: t > T0). Как видим из графика, (для данного примера) оптимальное "время жизни" бройлера на ~ 30% больше времени набора им половины предельного веса, а масса, при которой его отправляют в магазин равна ~ 72% от максимально достижимой (на рисунке масса дана в отношении к её максимальному значению, а масштабы цены и прибыли даны произвольно). Графики строились для таких численных параметров птицефабрики (без указания размерностей параметров, которые были даны): s = 10; а = 50; а = 0.5; β = 0.2. Ещё одно "открытие" полученное из модели: для малых: t ~ %·Τι/2 прибыль имеет... минимум.
Данная модель применима для расчёта параметров других аналогичных рынков, в частности для мясного животноводства, рыбного хозяйства и пр. Получили очень неожиданный в словесной интерпретации результат, что наибольшую валовую прибыль фабрика получает при реализации бройлеров по минимальной, но одновременно и по... оптимальной цене. Уравнение оптимума справедливо для любых зависимостей роста массы бройлеров: m(t).
Например, если скорость привеса некоторых объектов экспоненциально падает, или их масса экспоненциально насыщается от значения: т0 до (в пределе): т°°, то для m(t) имеем простое выражение: m(t) = m°° - (m°° - m0)*Exp(-t/C). Оптимизация прибыли для этого вида прироста массы дана на Рис. 2.32 (справа). Здесь оптимальная масса объекта ниже и равна: ~ 25% от её предельного значения и "время жизни" лишь ~ 40% времени набора им половины возможного (среднего у объектов данного типа) предельного веса. Саму кривую зависимости массы птицы от времени можно найти-построить экспериментально по точкам, по среднему значению массы птиц в контрольной группе, а затем полученные данные аппроксимировать подходящим аналитическим выражением m(t), с которым в дальнейшем можно проводить вышеуказанные аналитические операции поиска оптимума "времени жизни" и цены.
Но вернёмся к бройлерам и рассмотрим случай потери птицы в процессе их выращивания (травмы, несовместимые с жизнью, случайные не эпидемические смертельные заболевания). В этом трагическом случае число птиц убывает, и, как известно из анализа, убывание идёт по экспоненциальному закону, т.е., если ежедневно приобретается: Ко цыплят, то к моменту: t их останется: K(t) = Ko*Exp(-A*t), где параметр: λ полагается известным. Находим баланс затрат.
Затраты на ежедневное приобретение цыплят, как и прежде тривиальны, это: Qi = K0*s.
Затраты на содержание фабрики пропорциональны количеству бройлеров на ней в момент Т, а это количество равно: JK(t)dt = (Κο/λ)·[1 - Exp(-λ·T)]. Окончательно для затрат Ch имеем выражение: Q2 = α·(Κ0/λ)·[1 - Εχρ(-λ·Τ)], где коэффициент: а был описан выше. Что касается биомассы всех птиц, то она с учётом потерь будет: Ms = I K(t)»m(t)»dt, где интегрирование по t ведётся в пределах: {0...Т}. Поскольку: K(t) и m(t) известны, то интеграл вычисляется, и затраты на корм будут: Q3 = p*Ms. Уравнение связи параметров модели, при потерях в поголовье кур будет: р»Ехр(-х/а) = К(Т)»т(Т), откуда: K0 = [μ/ιη(Τ)]·Εχρ(λ·Τ - х/а). И прибыль фабрики: Q(x, Т) = р»х»Ехр(-х/а) - K0*s - α·(Κ0/λ)·[1 - Εχρ(-λ·Τ)] - β·Μδ. Вынесем за скобки параметр K0, и, проведя несложные преобразования, окончательно получим: Q(x, Т) = р»х»Ехр(-х/а) - K0*{s + (α/λ)·[1 - Εχρ(-λ·Τ)] + β-J m(t)*Exp(-A*t)*dt}. Дифференцируя по х, и приравняв производную к нулю, можно найти оптимальную отпускную цену мяса бройлеров: х(Т) = a + {s + (α/λ)·[1 - Εχρ(-λ·Τ)] + β·|m»Exp(-A*t)*dt}*Exp(A*t)/m(T). При λ => 0, эта формула совпадает с предыдущей для: х(Т), как это при отсутствии потерь птицы. На Рис. 2.32-1 даны те же графики, но при наличии потерь: А = 0.01352 (параметр взят таким, чтобы за 30 дней потери птицы составляли ~ 33.33%), и здесь мы видим, что...
наибольшая прибыль имеет место при минимальной цене бройлеров. Аналитическое же доказательство того, что и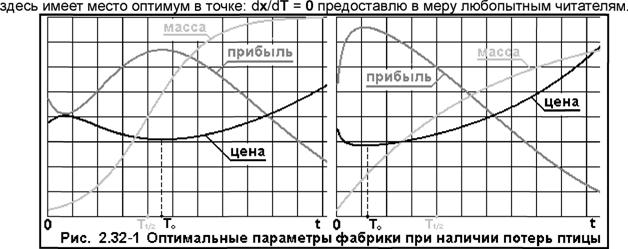 Как видим, при наличии потерь птицы, оптимальные цены (естественно) растут, но не так сильно, как потери птицы; прибыль падает, и "время жизни" птиц тоже сокращается. В нулевом приближении нами было принято, что затраты кормов пропорциональны массе птицы. А так ли это на самом деле? Рассмотрим проблему поподробнее. На что идёт питание |
вообще? Я, увы, не биолог, но дам свои соображения. Во-первых, часть еды идёт на чистый обмен веществ, - клетки выделяют шлаки-радикалы и требуют взамен новый материал. Во- вторых, часть пищи идёт на создание новых клеток, а в нашей задаче, - на наращивание массы птицы. И в-третьих, часть пищи идёт на "энергетику": на сохранение температуры тела, на поддержание пищеварения, на чисто механическое перемещение в пространстве и на т.п. служебные функции, не имеющие отношения к росту массы объекта. Ясно, что только первая часть пищи, (идущая на обменные процессы и на их "энергетику") и, частично, третья часть пропорциональны массе тела. Вторая часть (на "строительство") - пропорциональна скорости роста массы, а третья часть (энергетическая, идущая только на тепловыделение) пропорциональна площади поверхности тела, или массе в степени: 2/3. Поэтому, скорость потребления пищи любым организмом: q, можно представить в виде суммы трёх слагаемых: q = g»m + m' + смп2/3, где: д, с - это коэффициенты соответствующей размерности, которые в некотором приближении могут быть найдены из калорийности пищи, её состава (углеводы- белки-жиры), а также из неких биологических соображений-пропорций. Зная вид функции роста массы: m(t) = А/[В + Exp(-t/C)], можно получить динамику потребления пищи: q(t), как функцию времени. Всю пищу [кг]: Mn, требуемую на выращивание одной птицы в интервале времени: {0...Т}, найдём интегрированием динамики: q(t) по этому интервалу. После простых преобразований получим: Mn = Jq(t)»dt ξ g»Jm»dt +.fm'»dt + c*jm2/3*dt, где слагаемые будут:
- jm*dt = A*(C/B)*Ln{[B*Exp(+T/C) + 1]/(B + 1)]};
- jm'*dt = A*[Exp(+T/C) - 1]/[B«Exp(+T/C) + 1]/(B + 1);
- jm2/3»dt = 0(Α/Β)2/3·[Φ(χ2) - Φ(χι)], где: X2 = (1/B + 1)1/3; Xi = [1 + Ехр(-Т/С)/В]1/3; где принято: Φ(ζ) = 0.5·Ι_η[(ζ - 1)3/(ζ3 - 1)] - 305»Arctg[(2»z + 1)/305)]. Вид выражений, конечно, удручающий, но у них есть преимущество в том, что это окончательные решения в радикалах с известными параметрами, и их вычисление (при графических построениях) трудностей не составит.
Естественно, не факт, что сами коэффициенты: g и с постоянны во времени, но в первом приближении этой зависимостью пренебрежём. Сделаем еще интуитивное предположение, что в точке перегиба кривой: m(t) все три слагаемых: g»m, m' и с»т:/3 ~ одинаковы, или, что в период наибольшего роста пища идёт на всё поровну. Получим: m'(Ti/2) = 0.25»А/(В»С); m(Ti/2) = 0.5·(Α/Β); m2/3(Ti/2) = (0.5·Α/Β)23. Умножив последние уравнения на коэффициенты: (д и с) и приравняв их друг к другу, получим формулы для коэффициентов: g и с, а именно: g = 0.5/С, и с = (0.5/С)*(0.5*А/В)1/3, Итого, масса корма на птицу будет: Mn = (A/B/2)*Ln{[B*Exp(T/C) + 1]/ (В + 1)]} + А*[Ехр(Т/С) - 1]/[В*Exp(TVC) + 1]/(В + 1) + (А/В)-2 4/3 *[Ф(х2) - Ф(х·,)], а ежедневные затраты фабрики на корма будут: Q3 = {p*m(Ti/2)/[3»m'(Ti/2)]}»KMn ξ {%·β·0}·Κ·Μπ. Здесь в фигурных скобках {·} дано выражение цены корма, но через коэффициент: β, равный затратам на [кг] веса птицы. Остальные затраты-прибыли: (Qj) - не изменятся. Выражение для прибыли: Q(x, Т) = р»Ехр(-х/а)*[х - (s + α·Τ + 3/4»р»С»Мп)/т(Т)], а оптимальная цена будет: х(Т) = a + (s + α·Τ + %»p»C»Mn)/m(T). На Рис. 2.32-2 слева приведен графический способ поиска оптимальных параметров, аналогичный расчёту приведенному на Рис. 2.32, справа -
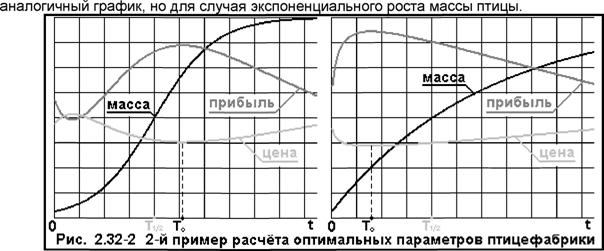 Как видим, в обоих моделях оптимальное "время жизни" птицы не изменилось, но для последней модели выросли цены (в точках оптимума), и, соответственно, снизилась прибыль. Варианты для случая потерь птицы предоставлю рассмотреть читателям самостоятельно. |
2.29.
Еще по теме Модель рынка бройлеров:
- Модели "чёрного" рынка и рынка наркоторговли
- Модель денежного рынка
- Модель фондового рынка.
- Модель рынка инвестиций
- 2.17. Модель рынка бартерного обмена
- Модель рынка монополиста-продавца
- Модель рынка монополиста-покупателя
- 4.3 Анализ динамики рынка, модель жизненного цикла
- Модель рынка скоропортящихся товаров
- Модель рынка аукционной торговли
- Модель конкурентного рынка продавцов
- Модель инфляционного рынка