6. 1.Основные постулаты
Эволюция — это процесс преобразования таксонов. Биологические таксоны представляют собой в большей или меньшей степени неперекрываю- щиеся (дискретные) группы фенотипов: каждый фенотип может принадлежать только одному таксону.
Благодаря своей дискретности таксоны отличаются друг от друга.Таксоны дискретны, потому что составляющие их фенотипы устойчивы по отношению к изменениям, постоянно происходящим в геноме популяций и в среде их обитания (при условии, что эти изменения не превышают определенного порогового значения). Следовательно, теория эволюции должна объяснять, каким образом происходит переход из одного устойчивого фенотипического состояния в другое (Шишкин, 1987, 1988а, 19886).
Устойчивость траектории развития, ведущей от зиготы к дикому типу, обозначается термином эквифипальность (по Г. Дришу) и отражает тот факт, что развивающийся организм может достичь одного и того же конечного состояния различными путями и следуя из различных начальных состояний (Webster, Goodwin, 1982). Это же свойство онтогенеза М. А. Шишкин (1987, 1988 а) называет забуференностью: развивающаяся система обладает множеством взаимосвязанных регуляторных механизмов, которые возвращают систему в прежнее состояние, если какие-либо факторы вызвали отклонение хода развития.
Устойчивая онтогенетическая траектория названа термином креод. В общем смысле креод — это траектория, которая притягивает близлежащие траектории (Уоддингтон, 1970). Зрительно креод может быть представлен в виде глубокого желоба, по которому скатывается шарик — развивающийся организм; если по какой-либо причине движение шарика отклоняется в сторону от дна желоба, то затем шарик вновь возвращается на стандартную траекторию.
Индивидуальное развитие способно создать не один, а множество фенотипов: креодам свойственно ветвиться. Зрительным образом такой системы разРис.
6. Эпигенетический ландшафт и повреждающие воздействия. А) Развитие переходит на дополнительный путь под влиянием внешнего шока (стрелка); Б) То же, под действием мутации, разрушившей стенку кре- ода; В) промежуточное состояние (по: Waddington, 1975; Шишкин, 1987, 1988 а).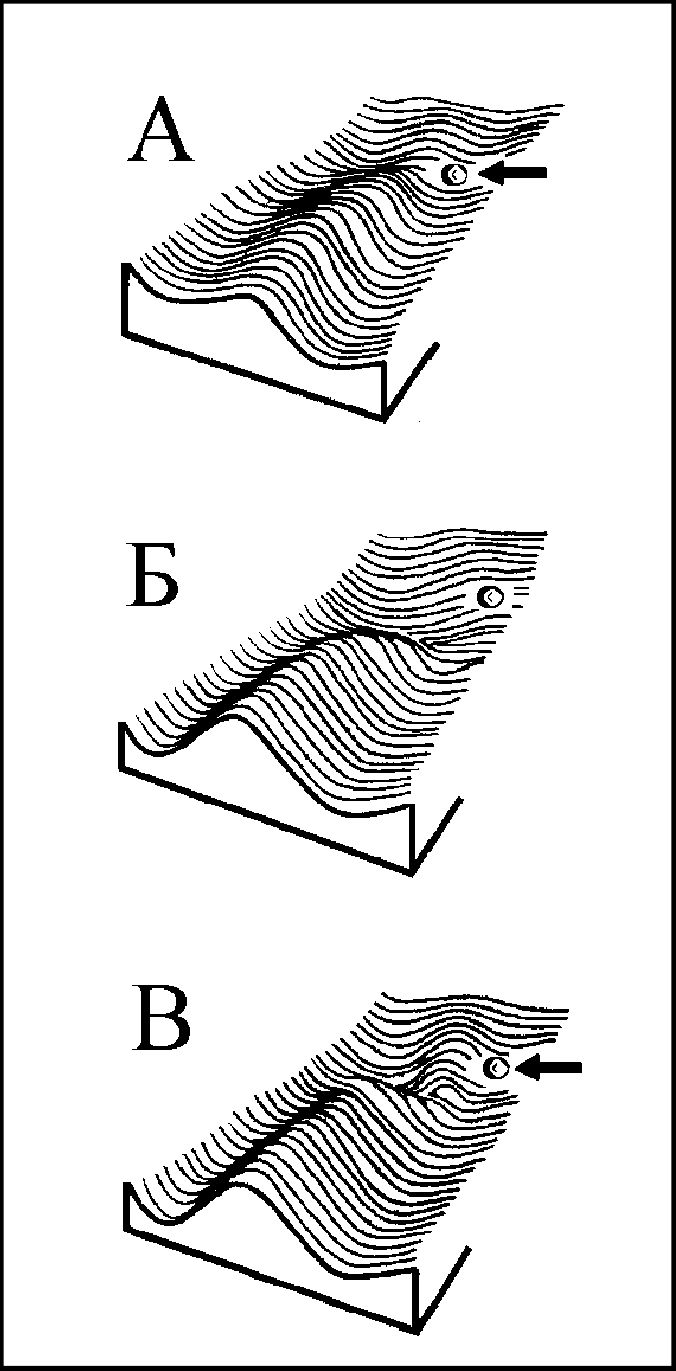
вития является эпигенетический ландшафт (Waddington, 1975)(рис. 6).
Развитие по основному креоду приводит к стандартному фенотипу, развитие по дополнительным траекториям — к онтогенетическим аберрациям, иначе называемым морфозами.
Морфозы — это необычные фенотипы, неустойчиво наследуемые уклонения онтогенеза, выходящие за пределы нормальной изменчивости данной популяции и возникающие в ответ на действие непривычных факторов (Шмальгаузен, 1946; Dobzhansky, 1951; Waddington, 1975; Се- верцов, 1987; Rieger et al., 1991). Морфозы могут также образовываться в ответ на селективно обусловленное изменение размеров тела (Alberch, 1981).
Морфозы возникают в результате нарушения зародышевых регуляционных механизмов. Признаки каждого морфоза детерминированы характером взаимодействия зародышевых зачатков и не зависят от природы факторов, вызывающих нарушение регуляции. Этот важнейший вывод, что морфологические признаки специфичны по отношению к состоянию зародыша и неспецифичны к характеру возмущений, был впервые сделан Р. Гольдшмидтом (Goldschmidt, 1938, 1940, 1955), который является, таким образом, родоначальником представления об онтогенезе как о системе из множества взаимосвязанных процессов, иначе — об эпигенетической системе.
Эпигенетикой называют раздел биологии о причинных взаимодействиях между генами и их продуктами, образующими фенотип (Уоддингтон, 1970), или иными словами, о механизмах онтогенетического развития (Hall, 1983; Lovtrup, 1988). Так или иначе, эпигенетика — синоним биологии развития или эмбриологии на клеточном уровне.
Эпигенетическая система есть нечто, находящееся над фенотипом и генотипом и определяющее их изменение в ходе эволюции.Нарушающие факторы могут быть внешними либо внутренними. Внешние факторы — это любые изменения условий существования и развития, настолько сильные, что вызывают переход развития на запасной креод (рис. 6 А). Внутренние факторы — это любые изменения структуры зиготы (в первую очередь — ее генотипа), изменяющие соотношение креодов, так что дополнительная траектория становится основной (рис. 6 Б). Можно сказать, что при действии внешнего толчка возникает фенокопия морфоза, а при наличии внутренних отклонений — его генокопия (на языке СТЭ — мутантный фенотип), причем резкой границы между фено- и генокопиями не существует, как не существует и ненаследуемых изменений: облик любого морфоза и вероятность его формирования в онтогенезе определяются структурой эпигенетического ландшафта; соответственно, любой морфоз в большей или меньшей мере наследуем (Шишкин, 19846).
С точки зрения различных теорий, фено- и генокопии имеют разное значение для эволюции. Для СТЭ главное — генокопии, возникающие при ре- комбинировании скрытых мутаций генов и хромосом. В соответствии с эпигенетической теорией естественный отбор оперирует преимущественно фенокопиями морфозов.
Эпигенетическая теория рассматривает эволюцию как процесс преобразования онтогенеза под влиянием изменений, происходящих в окружающей среде. Эта теория предполагает, что в ходе эволюционного изменения популяция старого вида вначале утрачивает фенотипическую устойчивость, и затем переходит в новое состояние, сперва очень изменчивое, но обретающее новую устойчивость в ходе последующей эволюции.
Начальная утрата устойчивости происходит под действием запорогового изменения в среде, где происходит индивидуальное развитие организма: смена условий развития нарушает нормальную координацию онтогенетических процессов, приводя к появлению значительного числа необычных фенотипов (фе- нокопий морфозов) и, соответственно, к возрастанию внутрипопуляционной изменчивости.
Фенокопии наследуются неустойчиво. Вместе с тем, они представляют новый материал для естественного отбора. Если какой-либо из вновь появившихся морфозов оказывается способным существовать в изменившихся условиях, то стабилизирующий отбор в пользу нового фенотипа сокращает изменчивость популяции и приводит к генетической ассимиляции этого морфоза (Waddington, 1975), перестраивая популяционный геном таким образом, что в пределах эпигенетического ландшафта происходит углубление одного из бывших запасных креодов. В результате фенокопия морфоза становится новой наследственной нормой, в соответствии с которой будет впредь проходить онтогенез большинства особей популяции. Последовательные утрата и приобретение фенотипической устойчивости составляют элементарный эволюционный цикл.Таким образом, эволюционное изменение начинается в экосистеме и заканчивается в геноме (Северцов, 1939; Красилов, 1984; Vermeij, 1987). Это положение делает эпигенетическую теорию альтернативой неодарвинизма, который рассматривает эволюцию как прямо обратную последовательность событий: от нового гена к новой экосистеме.
Еще по теме 6. 1.Основные постулаты:
- г) Основная норма как трансцендентально-логический постулат
- Основные постулаты развитой теории систем.
- Связь основных философских концепций исмаилизма с его религиозными постулатами. Рационализм философской доктрины исмаилизма – онтологический и гносеологический его планы. Место Бога в картине мироздания.
- VI. О постулатах чистого практического разума вообще
- § 2. Постулаты.
- 5.3. Постулаты Эйнштейна. Анализ понятий длины и времени. Преобразования координат
- К эвристическим постулатам причисляют следующие:
- Постулаты
- ПОСТУЛАТ
- ПОСТУЛАТ
- ПОСТУЛАТ
- 1. Постулаты этического биологизма
- § 6. Постулат де Лелека.
- § 6. Постулаты и истина
- Постулаты Экономической Таблицы
- Постулаты «практического разума»
- V. Бытие божье как постулат чистого практического разума
- 4. Постулаты эмпирического мышления вообще