IV.2. ХИМИЧЕСКОЕ СТРОЕНИЕ НУКЛЕИНОВЫХ КИСЛОТ
 Нукдеиновая кислота представляет собой гигантскую полимерную молекулу, построенную из многочисленных повторяющихся мономерных звеньев, называемых нуклеотидами.
Нукдеиновая кислота представляет собой гигантскую полимерную молекулу, построенную из многочисленных повторяющихся мономерных звеньев, называемых нуклеотидами.


Азотистые основания представляют собой производные одного из двух классов соединений — пуринового или пиримидинового ряда (рис. IV.З). В нуклеиновых кислотах присутствуют два производных пурина — аденин (обозначаемый А) и гуанин (Г) и три производных пиримидина — цитозин (Ц), тимин (Т) и урацил (У). В состав ДНК входят аденин, гуанин, цитозин и тимин. В структуре РНК тимин заменен другим пиримидином — урацилом.
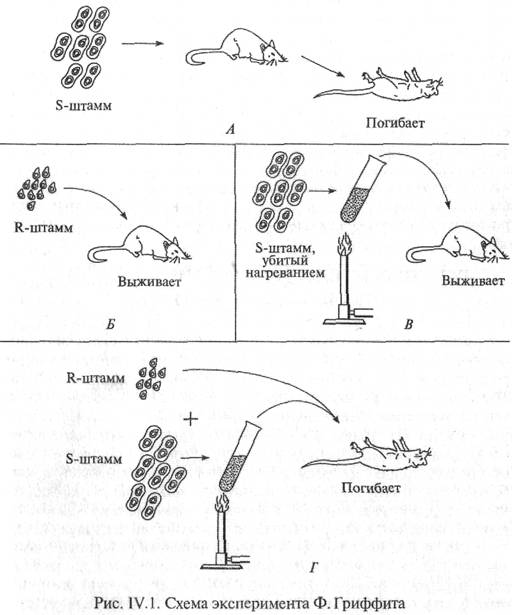
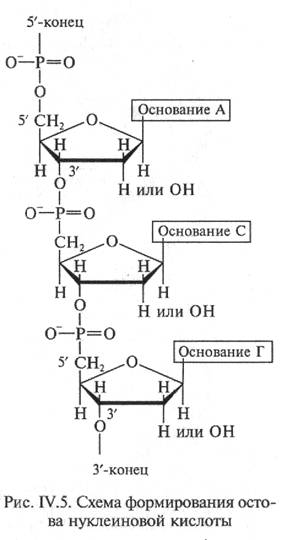
 | Сахар, входящий в состав нуклеотида, содержит пять углеродных атомов, т.е. представляет собой пентозу, которая может присутствовать в одной из двух форм: рибозы и дезоксирибозы (рис. IV.4). Различие между ними состоит в том, что атом водорода при втором атоме углерода дезоксирибозы в рибозе замешен гидроксильной группой (—ОН). Сахар присоединяется к одному из оснований с помощью гликозидной связи, соединяющей 1-й углеродный атом пентозы с 1-м атомом азота производных пиримидина или с 9-м атомом азота производных пурина. В зависимости от формы пентозы различают два типа нуклеиновых кислот - дезоксирибонуклеиновую кислоту (ДНК) и рибонуклеиновую кислоту (РНК). Нуклеиновые кислоты являются кислотами потому, что в их молекулу входит остаток фосфорной кислоты (-НРО3). В результате взаимодействия молекулы фосфорной кислоты с 3'-гидро-ксильной группой предшествующего ну-
|
клеотида и 5'-гидроксилъной группой после-ь дующего образуется основная цепь нуклеиновой кислоты (фосфат—сахар—фосфат—сахар и т.д.), к которой присоединяются разные азотистые основания (рис.
IV.5). Порядок следования оснований вдоль цепи носит название первичной структуры нуклеиновой кислоты. Последовательность оснований определяется в направлении от 5'- к З'-углеродному атому пентозы.
IV.2.1. Строение ДНК
| Пространственная конфигурация молекул ДНК была установлена в 1953 г. Это открытие было удостоено высшей научной награды — Нобелевской премии. Согласно модели, предложенной Дж.Уотсоном и Ф. Криком, молекула ДНК представляет собой две параллельные неразветвленные полинуклеотиднъте цепи, закрученные вокруг общей оси в двойную правостороннюю спираль. Диаметр двойной спирали равен 20 нм с шагом около 3,4 нм, на каждый виток приходится 10 пар нуклеотидов, поскольку расстояние между соседними парами составляет 0,34 нм (рис. IV.6). Направление цепей взаимно противоположно. На 5'-конце цепи ДНК всегда расположен свободный фосфат у 5'-атома углерода; на противоположном 3'-конце — свободная ОН-группа у З'-атома углерода. Сахарно-фосфатный остов располагается по периферии спирали, а азотистые основания находятся внутри ее. Между основаниями образуются специфические водородные связи (рис. IV.7). В 50-е годы было установлено, что в ДНК, выделенной из клеток различных организмов, молярная сумма пуриновых оснований (аденин + гуанин) всегда равна сумме пиримидиновьгх (цитозин + тимин). Содержание аденина равно содержанию тимина, а содержание гуанина — количеству цитозина (правило Э.Чаргаффа). | 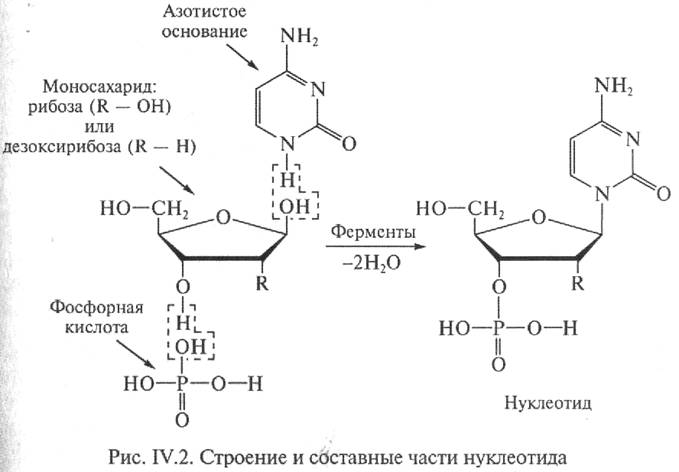 | |
 |  | |
Водородные связи возникают между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи в результате избирательного спаривания оснований.
Аденин всегда образует водородные связи с тимином (А-Т), а гуанин с цитозином (Г-Ц). Правило образования двунитевой структуры ДНК (А-Т, Г-Ц) называется правилом комплементарности (лат. complementum — дополнение). Образование водородных связей между комплементарными парами обусловлено их пространственным соответствием (рис. IV.8). Это приводит к тому, что нуклеотидные последовательности двух антилараллельных цепей ДНК всегда строго комплементарны друг к другу, а порядок чередования нуклеотидов в обеих цепях ДНК оказывается взаимообусловленным. Именно комплементарностью определяется точное воспроизведение последовательности оснований при копировании (репликации) молекул ДНК.Водородные связи между парами комплементарных нуклеотидов (две для пары А-Т и три для пары Г-Ц) относительно непрочные. Поэтому комплементарные нити молекулы ДНК могут разделяться и соединяться вновь при изменении некоторых условий (например, изменении температуры или концентрации солей). Разделение двухцепочечной ДНК называется денатурацией, а обратный процесс - образование двухцепочечной структуры ДНК - гибридизацией Очевидно, что в реакцию гибридизации могут вступать только комплементарные друг другу нуклеотидные последовательности одноцепочечных молекул.
Цепь, содержащая информацию о строении белка (в направлении 5'-3'), называется смысловой цепью, а комплементарная - антисмысловой. Антисмысловая цепь имеет большое значение при стабилизации структуры двойной спирали ДНК и участвует в процессах репликации и репарации (восстановления) поврежденных участков ДНК. Молекулы ДНК являются гигантскими полимерами. Единицами измерения длины молекулы приняты: пары нуклеотидов (п.н.), тысячи пар нуклеотидов - килобазы (кб), миллионы пар оснований - мегабазы (мб).
У человека гаплоидный набор содержит 3,2х109 пар нуклеотидов, или 3,2 миллиарда пар оснований. Почти вся ДНК клетки содержится в ядре в виде 46 плотно упакованных, суперскрученных за счет взаимодействий с ядерными белками, структурах - хромосомах. Сравнительно небольшая часть ДНК (около 5%) локализована в митохондриях.
Еще по теме IV.2. ХИМИЧЕСКОЕ СТРОЕНИЕ НУКЛЕИНОВЫХ КИСЛОТ:
- 1.1. Природные полисахариды хитин и хитозан: строение, физико-химические свойства
- Помимо усилий по выявлению необходимых благоприятных условий для химической эволюции, создаются теории, в которых объясняются варианты химической эволюции при возможных неблагоприятных условия
- 8.2. Определение выхода гуминовых кислот
- Вредные факторы производственной среды химического происхождения Источники вредных факторов химического происхождения на производстве
- Технология получения витамина С (L-аскорбиновая кислота)
- 8.2.2. Определение выхода свободных гуминовых кислот
- Совместное определение метилцеллозольва (этилцеллозольва) и метоксиуксусной (этоксиуксусной) кислоты
- Определение гликолевой кислоты в биологических объектах методом газовой хроматографии
- 8.2.1. Определение выхода общих гуминовых кислот
- ДНК (дезоксирибонуклеиновая кислота)
- Нарушения водно-электролитного баланса и кислотно-основного состояния.
- Физико-химическое действие ионизирующего излучения
- Определения целлозольвов и их метаболитов, а также гликолевой кислоты в биологических жидкостях
- Химически опасные объекты (ХОО)
- ИЗМЕНЕНИЕ СОДЕРЖАНИЯ НУКЛЕИНОВЫХ КИСЛОТ В ТКАНЯХ КАРПА В ЗАВИСИМОСТИ ОТ КОНЦЕНТРАЦИИ АЛЛОХТОННОГО АЗОТА В ВОДЕ И ТЕМПА РОСТА РЫБ