О ГЕНЕТИЧЕСКОМ И МОРФОЛОГИЧЕСКОМ РАЗНООБРАЗИИ MYTILUS EDULIS КАНДАЛАКШСКОГО ЗАЛИВА БЕЛОГО МОРЯ
Е.А. Жуковская
Московский государственный университет им. М. В. Ломоносова, Москва, Россия e-mail: zhukovskaya@mail.ru Введение
В начале 70- х годов появляются работы, посвященные изучению биоразнообразия и популяционной структуры моллюсков рода Mytilus, являющихся традиционным объектом промысла и культивирования, имеющих широкое распространение и играющих важную роль в морских экосистемах.
В основу большинства работ было положено широкое использование электрофоретически выявляемых полиморфных ферментных локусов, частоты аллелей и генотипы которых могут служить генетическими маркерами при изучении организации популяционной структуры и процессов происходящих внутри популяции (Tracey et al., 1975; Gosling, 1979; Koehn et al., 1984; McDonald, Siebenaller, 1989). В настоящее время доказано, что антропогенное влияние на окружающую среду приводит к разрушению биоценозов, к сокращению внутривидового разнообразия и к обеднению генофонда вида (Алтухов, 2003) Для выявления необратимых процессов в природе необходим постоянный мониторинг генетического и морфологического разнообразия природных скоплений. В задачу данной работы входило исследование генетического и морфологического разнообразия поселений Mytilus edulis Кандалакшского залива на основе использования биохимико-генетических маркеров.Материал и методы
Материалом для исследования послужили 7 выборок беломорских мидий (Mytilus edulis L.), собранные из естественных поселений в июле 1991 года, с разных участков литорали и верхней сублиторали Кандалакшского залива: 1 - пос. Лувеньга; 2 - Нефтебаза (ж/д ст. Белое море); 3 - губа Ковда; 4 - Кислая губа, кут (окрестности ББС МГУ), 5 - Круглая бухта, губа Чупа; 6 - пролив Оборина Салма (окрестности МБС СПбГУ), 7 - о. Соностров. Исследование генетического разнообразия проводили на основании распределения частот аллелей полиморфных локусов лейцинаминопептидазы (Lap) и неспецифических эстераз (Est), полученных при помощи метода электрофореза в полиакриламидном геле с системой прерывистых буферов.
В основу морфологической обработки материала были положены общепринятые морфологические измерения, используемые обычно при описании и идентификации раковин (Невесская,1965; Скарлато и Старобогатов,1979, McDonald et al., 1991) и на их основе рассчитаны индексы отношения: высоты створки к её длине (Н/L); выпуклости створки к её длине (Con/L); веса створки к произведению её длины, высоты и выпуклости (P/LxHxCon); длины аддуктора к длине ретрактора (La/Lr); длины ретрактора к длине створки (Lr/L); длины аддуктора к его ширине (La/Wa); ширины ретрактора к его длине (Wr/Lr). По каждому индексу вычисляли основные статистические параметры: среднее значение (М), ошибку средней (m), среднее квадратическое отклонение (σ), коэффициент вариации (С .V.). Статистическое сравнение выборок проводили с применением критериев Стьюдента (td), Фишера с использованием φ- преобразования, а также Пирсона (χ2).Результаты и обсуждение
Сравнительный анализ электрофореграмм показал, что по каждому локусу во всех выборках распределение генотипов соответствует закону Харди-Вайнберга. Частоты аллелей обоих локусов для каждой выборки приведены в таблице 1.
Таблица 1
Распределение частот аллелей исследованных локусов Lap и Est в выборках M.edulis
| № выборк и | Частота аллеля | |||||||||||
| Lap | Est | |||||||||||
| N | A | B | C | D | E | χ2 | N | A | B | C | χ2 | |
| 1 | 80 | 0.025 | 0.325 | 0.475 | 0.175 | - | 1 .29 | 76 | 0.158 | 0.553 | 0.289 | 1 .67 |
| 2 | 70 | 0.043 | 0.364 | 0.436 | 0.157 | - | 3.16 | 70 | 0.500 | 0.464 | 0.036 | 1 .61 |
| 3 | 1 50 | 0.010 | 0.253 | 0.590 | 0.147 | - | 12.39 | 80 | 0.044 | 0.600 | 0.356 | 2.89 |
| 4 | 80 | 0.012 | 0.188 | 0.625 | 0.175 | - | 2.41 | 52 | 0.250 | 0.317 | 0.433 | 2.86 |
| 5 | 1 45 | 0.028 | 0.297 | 0.500 | 0.155 | 0.020 | 5.84 | 61 | 0.213 | 0.533 | 0.254 | 6.72 |
| 6 | 70 | 0.036 | 0.371 | 0.429 | 0.114 | 0.050 | 3.09 | 70 | 0.278 | 0.529 | 0.193 | 5.41 |
| 7 | 64 | 0.047 | 0.328 | 0.398 | 0.164 | 0.063 | 3.01 | 64 | 0.218 | 0.508 | 0.274 | 1 .1 3 |
Примечание.
Обозначение выборок см. материал и методы.iopz2o.95(i=10)=18·3
Estx ..ц 3) =7.84
Как видно из приведенных данных распределение частот аллелей обоих локусов неравномерно в каждой выборке. По локусу Lap ведущим является аллель С. Его частота по выборкам колеблется от 0.398 до 0.625 при средней 0.493, а частота аллеля В от 0.188 до 0.371 при средней 0.304. Частота остальных аллелей этого локуса значительно меньше. Их средние составляют: А - 0.029, D - 0.155, E - 0.046. По локусу Est ведущим является аллель В с частотой колебания от 0.317 до 0.600 при средней 0.500, а частоты аллелей А и С колеблются в пределах 0.044 - 0.500 при средней 0.237 и 0.036 - 0.433 при средней 0.262. Необходимо отметить, что редкий аллель Е локуса Lap встречался только у трех выборок № 5, 6, 7 собранных в районе губы Чупа. Попарное сравнение выборок по частотам аллелей выявило ряд достоверных различий между ними (табл.2). В среднем каждая выборка достоверно отличается от 5 других по частоте встречаемости какого-либо аллеля.
Таблица 2
Сравнение выборок M.edulis по частотам аллелей исследованных локусов Lap и Est (критерий Фишера
с использованием φ - преобразования)
| № выборки | 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| 1 | * | --+-- | -++-- | --- + | --- + | --- + | ||
| 2 | +-+ | * | +++-- | -++-- | --- + | --- + | --- + | |
| 3 | +-- | +++ | * | --+-+ | -++-+ | +-+-+ | ||
| 4 | -++ | +++ | ++- | * | -++-+ | -++-+ | -++-+ | |
| 5 | --- | +-+ | +-+ | -++ | * | --- + | Lap | |
| 6 | +-+ | +-+ | +-+ | -++ | --- | * | ||
| 7 | --- | +-+ | +-- | -++ | --- | --- | * | |
| Est | ||||||||
Примечание. Сравнение частот аллелей A, B, C, D, E по исследованным локусам.
Наличие достоверных различий отмечено знаком +, отсутствие - .На рис. 1 приводится распределение выборок по частотам аллелей C обоих локусов в системе двух координат. Из приведенных данных видно, что выборки образуют три группы. Различия между группами столь велики, что даже доверительные интервалы с вероятностью 0.95 не перекрываются. В I группу входят выборки № 3 и № 4. Эти выборки достоверно не различаются между собой по частотам аллелей C исследованных локусов Lap и Est, но с высокой степенью достоверности, практически без перекрывания доверительных интервалов, отличаются от выборок остальных групп. Во II группу вошли четыре выборки (№ 1, 5, 6, 7), которые без перекрывания интервалов отличаются от выборок I группы по частотам аллелей C обоих локусов, а также отличаются от III группы по частоте аллеля C локуса Est. Несмотря на ряд достоверных различий, доверительные интервалы выборок этой группы перекрываются и поэтому нельзя в этой группе выделить по данным локусам какие либо подгруппы. В III группу вошла выборка № 2, отличающаяся с высокой степенью достоверности, без перекрывания доверительных интервалов, по частотам аллелей C обоих локусов от выборок I группы и по частоте аллеля C локуса Est от выборок II группы.
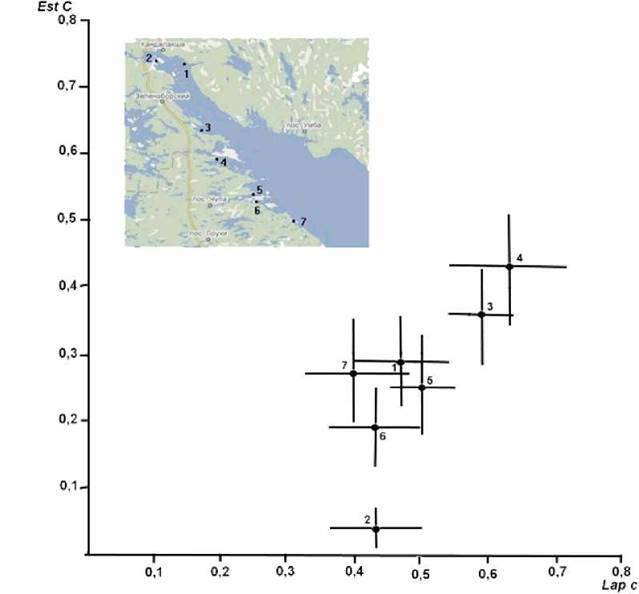
Рис.1. Распределение выборок в системе координат по частотам аллелей исследованных локусов. Точка - частота аллеля в выборке, тонкие линии - доверительные интервалы при р = 0,05. На карте Кандалакшского залива места сбора выборок отмечены точками. 1 - пос. Лувеньга; 2 - Нефтебаза (ж/д ст. Белое море); 3 - губа Ковда; 4 - Кислая губа, кут (окрестности ББС МГУ), 5 - Круглая бухта, губа Чупа; 6 - пролив Оборина Салма (окрестности МБС СПбГУ), 7 - о. Соностров
Таблица 3
Результаты статистического сравнения выборок по морфологическим индексам (критерий Стьюдента td)
| № выборки | 2 | 3 | 4 | 5 | 6 | 7 |
| 1 | -++-+-+ | +++++-+ | +-+-+++ | -++---+ | +++++++ | |
| 2 | * | -+++-+- | +-+-+-+ | +++-+-+ | -++-+-+ | +++-+-+ |
| 3 | * | +++++-+ | +-+-+++ | +++-+++ | -++++++ | |
| 4 | * | ++++-+- | +++++-- | -++-+++ | ||
| 5 | * | +++---- | ++++--+ | |||
| 6 | * | ++++--+ |
Примечание.
Расположение сравнений морфологических индексов в ячейке: H/L, Con/L, P/(L*H*Con), La/Lr, Lr/L, La/Wa, Wr/Lr.Наличие достоверных отличий отмечено знаком +, отсутствие -. Обозначение выборок см. на рис.1.
В таблице 3 представлены результаты статистического сравнения выборок по семи морфологическим индексам. Анализ результатов попарного сравнения выборок по морфологическим индексам показал чрезвычайную вариабельность выборок, определяющих форму раковины. Необходимо отметить, что выборка № 1 из пос. Лувеньга не имела достоверных различий ни по одному морфологическому индексу с выборкой № 3 из губы Ковда, но достоверно отличалась по семи морфологическим индексам от Соностровской выборки № 7 и по шести от выборки № 4 из Кислой губы. Ка- 223
ждая выборка в среднем отличается от другой по 5 морфологическим индексам. Выборки, собранные из района губы Чупа (№ 5, № 6 и № 7) не имели достоверных различий по индексам Lr/L, La/Wa, отражающих форму аддуктора и ретрактора. Наиболее вариабельным оказался признак P/(L*H*Con) по средним значениям которого достоверно различались все выборки и Con/L, по которому достоверно различалось большинство выборок.
Выводы
Таким образом, проведенный по биохимико- генетическим признакам анализ показал генетическую неоднородность локальных поселений мидий Mytilus edulis Кандалакшского залива и выявил её структурированность в виде трех дискретно различающихся групп выборок. На основании полученных данных пока трудно диагностировать причину различий между группами. С одной стороны она может быть сугубо генетической, связанной с ослаблением миграции личинок между поселениями мидий. С другой стороны различия между группами могут быть связаны с различной экологией участков, с которых брали пробы. Кроме того, возможно совместное действие этих факторов. Для выяснения этого вопроса требуются дальнейшие биохимико - генетические исследования с параллельным изучением экологических условий залива.
Анализ морфологического разнообразия показал большую вариабельность локальных поселений мидий по морфологическим индексам.
Наиболее изменчивым были индексы отношения: веса створки к произведению её длины, высоты и выпуклости (P/LxHxCon), и выпуклости створки к её длине (Con/L). Столь значительное разнообразие выборок по форме раковин определяется, очевидно, различными экологическими условиями в местах обитания мидий.Литература
Алтухов Ю.П., 2003. Генетические процессы в популяциях. М.: ИКЦ «Академкнига». 431 с.
Невесская Л.А., 1965. Позднечетвертичные двустворчатые моллюски Черного моря, их систематика и экология. М.: Наука. Т. 105. 386 с.
Скарлато О.А., Старобогатов Я.И., 1979. Положение в систематике и распространение мидий // Промысловые двустворчатые моллюски-мидии и их роль в экосистемах. Л.: Зоол. ин-т. С. 106-111.
Gosling E., 1979. Hidden genetic variability in two populations of a marine mussel // Nature. V. 279. № 5715. P. 713-715.
Koehn R.K., Hall G.J., Jnnes D.J., Zera A.J., 1984. Genetic differentiation of M.edulis in eastern North America // Marine Biol. V. 79. P. 117-126.
McDonald J.H., Siebenaller A., 1989. Similar geographic variation at the Lap locus in the mussel Mytilus trossulus and M.edulis // Evolution. V. 43. № 1. P. 228-231.
McDonald J.H., Seed R., Koehn R.K., 1991. Allozymes and morphometric characters of three species of Mytilus in the Nothern and Southern Hemispheres // Mar.Biol. V. 111. P. 323-333.
Tracey M.L., Bellet N.F., Gravem C.D., 1975. Excess allozyme homozygosity and breeding structure in the mussel Mytilus californianus // Mar.Biol. V. 32. № 3. P. 303-311.
ON GENETICAL AND MORPHOLOGICAL DIVERSITY OF MYTILUS EDULIS IN THE KANDALAKSHA BAY OF THE WHITE SEA E.A. Zhukovskaya
Biological Faculty, the Moscow State University (named after Lomonosov), Moscow, Russia
e-mail: zhukovskaya@mail.ru
The genetic and morphological variety of seven natural settlements of mussels Mytilus edulis from the Kandalaksha bay of the White Sea was studied. The analysis of the distribution of frequencies alleles polymorphic loci leucine aminopeptidase (Lap) and nonspecific esterase (Est), received by the electrophoresis in polyacrylamide gel, has shown genetic heterogeneity of local settlements of mussels in the Kandalaksha bay and has revealed its structure in the form of three discretely differing groups samples. It was shown that the samples are diverse in every studied morphological character and the degree of characters' variability in the samples is different. The most variable indexes were: weight to multiplication of length, height and convexity of a shell (P/LxHxCon), and convexity to length (Con/L). The given effect can be considered as the result of the influence of ecological conditions on growth and formation of the shells.
Еще по теме О ГЕНЕТИЧЕСКОМ И МОРФОЛОГИЧЕСКОМ РАЗНООБРАЗИИ MYTILUS EDULIS КАНДАЛАКШСКОГО ЗАЛИВА БЕЛОГО МОРЯ:
- ДИНАМИКА СТРУКТУРЫ ПОСЕЛЕНИЯ MYTILUS EDULIS НА ЛИТОРАЛИ КАНДАЛАКШСКОГО ЗАЛИВА БЕЛОГО МОРЯ
- СРАВНИТЕЛЬНОЕ СОДЕРЖАНИЕ РТУТИ В МЫШЦАХ РЫБ ВОДОЕМОВ СЕВЕРА ЕВРОПЕЙСКОЙ РОСИИ (КАНДАЛАКШСКИЙ ЗАЛИВ БЕЛОГО МОРЯ)
- ВЛИЯНИЕ ЛЕКТИНОВ РАЗЛИЧНОЙ УГЛЕВОДНОЙ СПЕЦИФИЧНОСТИ НА ГЕМОЛИТИЧЕСКУЮ АКТИВНОСТЬ ГЕМОЦИТОВ МИДИИ MYTILUS EDULIS
- ВОЗМОЖНОСТЬ ЗАВИСИМОСТИ ВАРЬИРОВАНИ СЕРДЕЧНОЙ АКТИВНОСТИ MYTILUS EDULIS L. И MODIOLUS MODIOLUS L. ОТ ФЛУКТУАЦИЙ ГИДРОЛОГИЧЕСКИХ ПОКАЗАТЕЛЕЙ
- О ЛЕТНЕМ ИХТИОПЛАНКТОНЕ ДВИНСКОГО ЗАЛИВА БЕЛОГО МОРЯ
- ВЛИЯНИЕ РАЗНОЙ СОЛЕНОСТИ НА ЛИПИДЫ АМФИПОД БЕЛОГО МОРЯ
- СОСТАВ КРИОФЛОРЫ ПРИБРЕЖНЫХ ЛЬДОВ КАНДАЛАКШСКОГО ЗАЛИВА БЕЛОГО МОРЯ
- К ЭКОЛОГИИ ПРИМОРСКИХ ОСОК (CAREX L) КАНДАЛАКШСКОГО ЗАЛИВА БЕЛОГО МОРЯ
- СТРУКТУРА И ДИНАМИКА ПРИМОРСКИХ РАСТИТЕЛЬНЫХ СООБЩЕСТВ ПОМОРСКОГО И КАРЕЛЬСКОГО БЕРЕГОВ БЕЛОГО МОРЯ Л.А. Сергиенко
- СЕЗОННО-ОБУСЛОВЛЕННЫЕ ИЗМЕНЕНИЯ В ТРАНСМИССИИ ПАРАЗИТОВ В ПРИБРЕЖЬЕ БЕЛОГО МОРЯ
- СОСТОЯНИЕ ГИДРОБИОЦЕНОЗОВ РЕК ПОМОРСКОГО ПОБЕРЕЖЬЯ БЕЛОГО МОРЯ
- КРИОФАУНА ЛЬДОВ ПРОЛИВА ВЕЛИКАЯ САЛМА КАНДАЛАКШСКОГО ЗАЛИВА БЕЛОГО МОРЯ
- МОРФОЛОГИЧЕСКИЕ АСПЕКТЫ ВИДООБРАЗОВАНИЯ И ФОРМИРОВАНИЯ АДАПТАЦИОННЫХ СТРАТЕГИЙ ВИДОВ Р. COCHLEARIA L. В ПРИБРЕЖНЫХ РАСТИТЕЛЬНЫХ СООБЩЕСТВАХ БЕЛОГО МОРЯ Л.А. Сергиенко
- РАЗМЕРНЫЙ СОСТАВ И ЛИПИДНЫЕ ПОКАЗАТЕЛИ У АМФИПОД БЕЛОГО МОРЯ В РАЗНЫХ УСЛОВИЯХ ОБИТАНИЯ Г.А. Шкляревич1, В.В. Богдан2
- ПРОДУКЦИОННЫЕ ХАРАКТЕРИСТИКИ ФИТОПЛАНКТОНА БЕЛОГО МОРЯ В ЗАВИСИМОСТИ ОТ ИСТОЧНИКА АЗОТА
- МОРФОБИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА СИГА (COREGONUS LAVARETUS L.) БАССЕЙНА ГУБЫ ЧУПА БЕЛОГО МОРЯ
- ПОЛОВАЯ СТРУКТУРА МОРСКОЙ ЗВЕЗДЫ ASTERIAS RUBENS L. НА ТЕРРИТОРИИ КАНДАЛАКШСКОГО ЗАЛИВА БЕЛОГО МОРЯ В 2003 ГОДУ
- НОВЫЕ ДАННЫЕ О ВОДНЫХ БЕСПОЗВОНОЧНЫХ БЕЛОГО МОРЯ В ПИТАНИИ ЛЕСНЫХ ПТИЦ
- БИОАККУМУЛЯЦИЯ ТЯЖЕЛЫХ МЕТАЛЛОВ РАЗЛИЧНЫМИ КОМПОНЕНТАМИ ЭКОСИСТЕМЫ КАНДАЛАКШСКОГО ЗАЛИВА БЕЛОГО МОРЯ
- ПЕРВИЧНАЯ ПРОДУКТИВНОСТЬ ФИТОПЛАНКТОНА В ПРОЛИВЕ ПЕЧАКОВСКАЯ САЛМА БЕЛОГО МОРЯ В ЛЕТНИЙ ПЕРИОД.