ГЕНЕТИЧЕСКАЯ СТРУКТУРА ПОПУЛЯЦИИ АТЛАНТИЧЕСКОГО ЛОСОСЯ (SALMO SALAR) БАССЕЙНА БЕЛОГО МОРЯ, ВЫЯВЛЕННАЯ С ПОМОЩЬЮ МИКРОСАТЕЛЛИТНЫХ МАРКЕРОВ[2]
А.К. Тонтери 1, А.Е. Веселов2, А.В. Зубченко3, Я.И. Лумме4, С.М. Калюжин5, М.Ю. Озеров1, К.Р. Приммер1
1Департамент биологии, подразделение генетики и физиологии, Университет Турку, Финляндия 2Учреждение Российской академии наук Институт Биологии Карельского научного центра РАН, г.
Петрозаводск, Россия.3Полярный НИИ морского рыбного хозяйства и океанографии им. Н.М. Книповича, г. Мурманск, Россия 4Департамент биологии, Университет Оулу, Финляндия.
5Варзугский научно-исследовательский центр полярных экосистем, с. Варзуга, Россия
e-mail: mikhail.ozerov@utu.fi
Введение
Популяции атлантического лосося, нерестящиеся в реках бассейна Белого моря, следует рассматривать как одни из немногих, не затронутых сильным антропогенным воздействием (Parrish et al., 1998). Лосось этого региона, являясь главным источником биологического разнообразия для всей Европы, также имеет огромное экономическое и социальное значение, проявляющееся, например, в виде рыболовного туризма. Однако, в настоящее время, знания о генетической структуре популяций атлантического лосося бассейна Белого моря основаны, в первую очередь, на изучении митохондриальной ДНК (Asplund et al., 2004), а так же на нескольких работах, с использованием ядерных маркеров, включающих лишь небольшую часть популяций региона (Kazakov, Titov, 1991; Makhrov et al., 2005; Saisa et al., 2005; Tonteri et al., 2005). Поэтому одной из основных задач данной работы было проведение более детального исследования генетической структуры популяций с использованием ядерных маркеров. Полученную с их помощью информацию необходимо использовать для разработки планов по сохранению и управлению генетическими ресурсами. Эта информация может послужить основой для идентификации принадлежности особей, выловленных в открытом море, к той или иной популяции.
Одним из глобальных природных явлений, оказавших значительное влияние на генетическую структуру атлантического лосося бассейна Белого моря, рассматривается ледниковый период. Массивный слой льда покрывал всю эту область тысячи лет, пока лед не отступил около 11,5-12 тыс. лет назад (Svendsen et al., 2004). Постледниковая история атлантического лосося, распространенного на северо-западе России, уже изучалась до этого, и были предложены несколько сценариев реколонизации. А.А. Махров с соавторами (Makhrov et al., 2005) предположил, что колонизация бассейна Белого моря происходила из бассейна Балтики вместе с мигрантами из Атлантического океана, а бассейн Баренцева моря был заселен как из восточной, так и из западной Атлантики. О балтийском влиянии в регионе также предположили Р.В. Казаков и С.Ф. Титов (Kazakov, Titov, 1991). Однако Т. Асплунд с соавторами (Asplund et al., 2004) предположили, что колонизация рек, впадающих в Белое и Баренцево моря, происходила напрямую из восточного рефугиума (убежища), т.е. лососем, нагуливающимся в Атлантическом океане и позже мигрировавшим на Кольский полуостров. Гипотеза восточного убежища и дополнительной миграции с запада была впоследствии поддержана в работе А. Тонтери с соавторами (Tonteri et al., 2005).
Целью данной работы заключалась в изучении генетической структуры популяций атлантического лосося с помошью ядерных маркеров. В связи с этим был проведен микросателлитный анализ большого числа популяций, воспроизводящихся в нерестовых реках бассейна Белого и Баренцева морей. Всего 1352 особи атлантического лосося, представленных 34 популяциями Северного, Норвежского, Баренцева и Белого морей, были изучены с использованием 14 микросателлитных маркеров (рис. 1). Дополнительно эти данные применили для оценки относительных пропорций общего генетического разнообразия региона, что может являться основой для разработки планов по управлению генетическими ресурсами и их сохранению. Наконец, была проверена правильность данных при использовании их для идентификации принадлежности особей, выловленных в зонах рыболовного промысла, к той или иной популяции.
Принадлежность к группам, выявленным с помощью анализа главных компонент, обозначена цветом: черный - Атлантика и Запад Баренцева моря, штриховка - Восток Баренцева моря, серый - Запад Белого моря, белый - Кольский п-ов. Реки: I-Dee - Ди, 2-Soy - Сёйя, 3-Rei - Рейса, 4-Ten - Тенойоки, 5-Tit - Титовка, 6-ZaL - Западная Лица, 7-Ura - Ура, 8-Tul - Тулома, 9-Kola - Кола, 10- Dro - Дроздовка, 11-Yok - Йоканьга, 12-Kac - Качковка, 13-Leb - Поной Лебяжья, 14-Pac - Поной Пача, 15-Dan - Даниловка, 16-Bab - Бабья, 17-Lih - Лиходеевка, 18-PuK - Пулоньга (Кольский п- ов), 19-Yug - Югина, 20-Kit - Кица, 21-Yap - Варзуга Япома, 22-Ole - Оленица, 23-Umb - Умба, 24- Pil - Пила, 25-Kolv - Колвица, 26-Nil -Нильма, 27-PuW - Пулоньга (Белое море), 28-Pon - Поньго- ма, 29-Sum - Сума, 30-Emt - Северная Двина Емца, 31-Pad - Северная Двина Падома, 32-Meg - Ме- гра, 33-Piz - Печора Пижма, 34-Unj - Печора Унья.
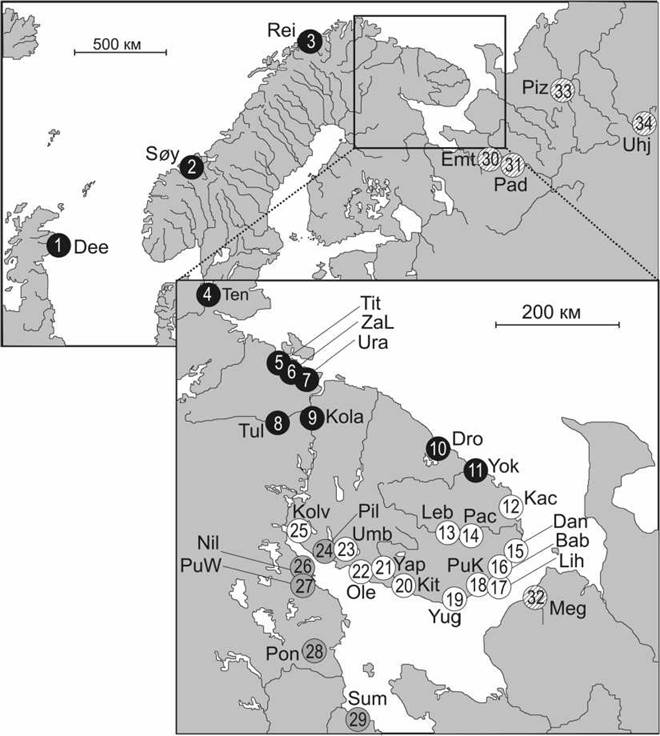
Рис. 1. Схема мест сбора образцов популяций атлантического лосося.
Результаты и обсуждение
На основании данных микросателлитной изменчивости исследованные популяции были разделены на четыре кластера. Популяции атлантического лосося, воспроизводящиеся в реках западной части Баренцева моря, образовали единый кластер с популяциями, нерестящимися в реках северовосточной Атлантики. Кроме того, популяции из рек Белого и восточной части Баренцева моря были разделены на три дополнительные группы (рис. 2). Популяции востока и юга Кольского полуострова (Белое море) составили одну группу, за исключением популяций из рек Оленица и Пила, которые попали в кластер с популяциями западного побережья Белого моря, формирующих третью группу. Четвертая группа была сформирована популяциями восточной части Белого моря и Баренцева моря за исключением популяции реки Мегра (Архангельская область), которая попала в группу с географически близкими популяциями восточной части Кольского полуострова.
Популяция реки Сума из южной части Белого моря оказалась в одной группе с популяциями из восточной части Белого и Баренцева морей. Сформированные кластеры четко соотносятся с географическими регионами отбора образцов. Наиболее вероятно, что полученные группы отражают историю колонизации региона. Вместе с тем, теоретически, границы между ними должны медленно исчезать, т.к. они не поддерживаются естественным отбором. Однако необходимы дальнейшие исследования, прежде чем могла бы быть определена относительная роль естественного отбора и филогеографической истории.Никакой разницы в уровне генетического разнообразия не было отмечено между группами Атлантики, западной части Баренцева моря и Кольского полуострова или между группами западной части Белого моря и восточной части Баренцева моря (табл.). Однако уровень генетического разнообразия популяций западной части Белого моря (Карелия) был достоверно ниже, чем популяций Атлантики, западной части Баренцева моря и популяций Кольского полуострова. Так же и популяции восточной части Баренцева моря показали более низкие значения генетического разнообразия, чем популяции Атлантики, западной части Баренцева моря и популяции Кольского полуострова.
Показатели генетического различия (FST) внутри групп для кластеров западной части Белого моря и восточной части Баренцева моря были достоверно выше по сравнению с двумя другими группами (Атлантики, западной части Баренцева моря и группы Кольского полуострова). Однако показатели FST для групп западной части Белого моря и восточной части Баренцева моря не отличались достоверно друг от друга, так же как и эти показатели для двух дру-
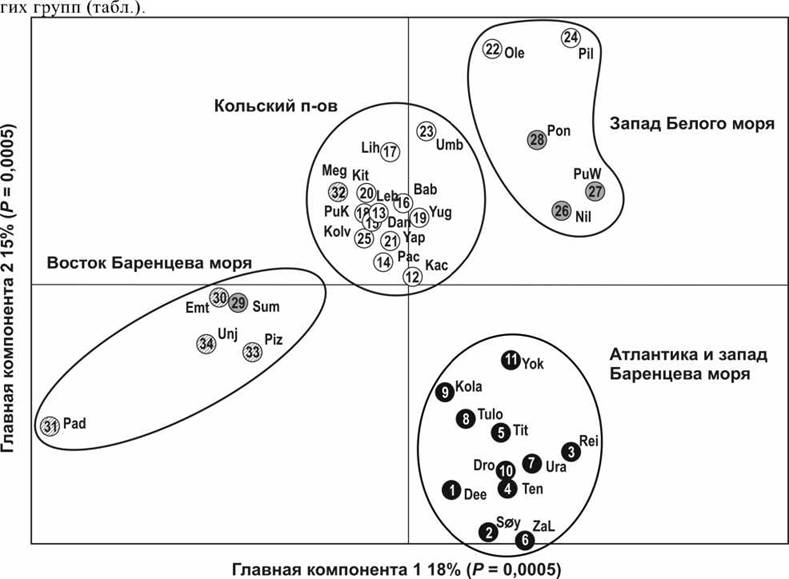
Рис. 2. Распределение лосося в пространстве главных компонент: черные кружки - популяции лосося северо-западной части Атлантического океана и западной части Баренцева моря, белые - восточной и южной частей Кольского п- ова, серые - западной части Белого моря, заштрихованные - восточной части Белого и Баренцева морей.
Успешность моделирования распределения выловленных на нагуле особей в свою речную популяцию сильно различалась. На уровне групп успех определения был относительно выше, показывая, что большинство неправильных определений на уровне популяции было опосредовано особями, попавшими в другую популяцию из той же группы (табл.). В некоторых случаях низкий уровень успешности являлся следствием слишком малого числа отобранных для исследований особей. Тем не менее, доля успешных попыток при определении региона происхождения была относительно высокой (87-96%), указывая, что этих данных было достаточно для относительно точного определения на таком уровне (табл.).
Средние показатели генетического разнообразия и различия популяций, выявление достоверного сигнала изоляции расстоянием внутри региона и средняя степень успешного распределения особей на уровне популяции и подгрупп
| Регион | Генетическое разнообразие (Ar, He) | Генетическое различие (FST) | Достоверность изоляции расстоянием? | Средняя степень успешного распределения (%): | |
| в подгруппу | в популяцию внутри подгруппы | ||||
| Запад Баренцева моря / Атлантика | 7,3; 0,73 | 0,032 | Да | 87 | 62 |
| Кольский п-ов (Белое море) | 7,1; 0,71 | 0,022 | Да | 93 | 45 |
| Запад Белого моря | 5,1; 0,65 | 0,101 | Нет | 96 | 95 |
| Восток Баренцева моря | 5,3; 0,64 | 0,108 | Нет | 96 | 90 |
Таким образом, с относительно высокой степенью точности полученные данные могут быть использованы для определения региона происхождения особей, которые выловлены в открытом море (рис.
2). Однако для идентификации на уровне популяций требуются исследования большего количества локусов и, в некоторых случаях, большего количества образцов.Изоляция расстоянием определялась путем попарных сравнений генетических дистанций, выраженных как FST/(1-FST) и географических расстояний, измеренных как кратчайший водный путь между местами взятия образцов (в км). Несмотря на то, что показатель изоляции расстоянием был достоверным при включении в анализ всех исследованных популяций, для отдельных групп достоверная изоляция расстоянием была отмечена только среди популяций образующих группу Кольского полуострова (бассейн Белого моря), группу Атлантики и западной части Баренцева моря, тогда как в других двух группах не было отмечено признаков изоляции расстоянием (рис. 3, табл.). Это указывает на то, что пространственный масштаб, на котором была изучена изоляция расстоянием, имеет значение, что также было отмечено и в более ранних работах (Slatkin, 1993; Rousset, 1997; Ehrich, Stenseth, 2001; Castric, Bernatchez, 2003). Нами
был обнаружен существенный разброс в наклоне линии регресса при изоляции расстоянием, когда рассматриваемые популяции были отдаленны друг от друга на расстояние 100 км или меньше.
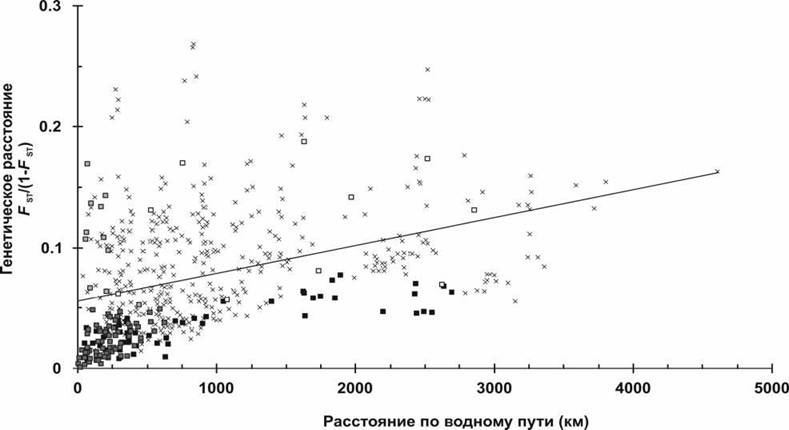
Рис. 3. Связь между географическими и генетическими расстояниями изученных популяций атлантического лосося. Квадратами обозначены межпопуляционные расстояния в каждой из групп, выявленные анализом основных компонент
(черные - Атлантика и западная часть Баренцева моря; темно серые - Кольский п-ов; светло серые - западная часть Белого моря; белые - восточная часть Баренцева моря), межпопуляционные расстояния между четырьмя группами представлены крестиками. Линия показывает регрессию всех межпопуляционных сравнений (rXY = 0,39; P = 0,008).
Наблюдаемая ситуация могла бы быть объяснена прохождением популяциями состояния «бутылочного горлышка», т.е. относительно короткого периода времени, когда в силу определенных, непредсказуемых экстремальных ситуаций выживает небольшое количество особей от начальной численности популяции. Вследствие этого теряется большая часть оригинального генетического разнообразия. Чаще всего «бутылочное горлышко» вызвано изменениями окружающей среды. Однако в нашей работе его не было выявлено. Наиболее вероятное объяснение состоит в том, что изученные популяции находятся в неравновесной ситуации по потоку/дрейфу генов, поскольку вся изученная область была скрыта массивным ледяным покровом в течение последнего ледникового максимума приблизительно 25-15 тыс. лет назад (Svendsen et al., 2004). Архангельская область была освобождена ото льда приблизительно 13 тыс. лет назад, а Белое море и большая часть Кольского полуострова приблизительно 11,5-12 тыс. лет назад (Svendsen et al., 2004). Таким образом, вероятно, атлантический лосось из Белого моря все еще проходит временную фазу перехода к равновесию, и сигнал изоляции расстоянием не может быть обнаружен на больших географических дистанциях.
Четкое разделение популяций на группы: побережье Белого моря, включая Кольский полуостров, северо-восточную Атлантику и западное побережье Баренцева моря, а также на восточное побережье Баренцева моря, указывает на то, что эти регионы были колонизированы из разных убежищ. Реки Белого моря и восточного побережья Баренцева моря были колонизированы с востока, как и предполагалось до этого (Asplund et al., 2004; Tonteri et al., 2005), и подтверждается настоящими наблюдениями различных характеристик для популяций этой области. Предположения о миграции с запада в западную часть Баренцева моря были сделаны в предыдущих исследованиях (Asplund et al., 2004; Makhrov et al., 2005; Tonteri et al., 2005), а так же подтверждаются результатами данной работы. Вышеизложенное также позволяет сделать заключение, что западная часть Баренцева моря была колонизирована скорее с запада, чем с востока после схода ледника, или, что западные линии, по крайней мере, значительно повлияли на генетический состав современных популяций атлантического лосося этой области.
Заключение
Так какие же популяции должны быть сохранены в первую очередь, согласно обнаруженной в регионе генетической структуре? Четыре группы с различной послеледниковой историей, выявленные в этой работе, могут рассматриваться в качестве отправной точки для определения четырех единиц управления генетическими ресурсами. Несомненно, для этих четырех групп могут потребоваться различные стратегии управления, поскольку были выявлены два различных типа генетического разнообразия и различия (табл.). Популяции лосося, нерестящиеся в реках Кольского полуострова в бассейне Белого моря, а также в бассейне Атлантического океана и западного побережья Баренцева моря показали большее генетическое разнообразие и меньшее генетическое различие, чем популяции воспроизводящиеся в реках западного побережья Белого моря и восточного побережья Баренцева моря. При этом у двух предшествующих групп наблюдается изоляция расстоянием (табл.). Это означает, что относительная значимость потока генов по сравнению с их дрейфом играет разную роль в определении структуры популяции в этих четырех регионах. Поток генов оказывает большее влияние в реках Кольского полуострова, Атлантики и западной части Баренцева моря, а дрейф генов - в западной части Белого моря и восточной части Баренцева моря.
Работа выполнена при финансовой поддержке РФФИ (грант №08-04-91771-АФ_а), Академии наук Финляндии (грант №124121), Программы фундаментальных исследований Президиума РАН «Биологическое разнообразие» (2009-2011 гг.) и Варзугского научно-исследовательского центра полярных экосистем.
Литература
Asplund T., Veselov A., Primmer C.R., Bakhmet I., Potutkin A., Titov S., Zubchenko A., Studenov I, Kaluzchin S., Lumme J., 2004. Geographical structure and postglacial history of mtDNA haplotype variation in Atlantic salmon (Salmo salar L.) among rivers of the White and Barents Sea basins // Annales Zoologi Fennici V. 41.
P. 465-475.
Castric V., Bernatchez L. 2003. The rise and fall of isolation by istance in the anadromous brook charr (Salvelinus fontinalis Mitchill) //Genetics V. 163. P. 983-996.
Ehrich D., Stenseth N.C. 2001. Genetic structure of Siberian lemmings (Lemmus sibiricus) in a continuous habitat: large patches rather than isolation by distance // Heredity V. 86. P. 716-730.
Kazakov R.V., Titov S.F. 1991. Geographical patterns in the population genetics of Atlantic salmon, Salmo
salar L., on U.S.S.R. territory, as evidence for colonization routes // Journal of Fish Biology V. 39. P. 1-6.
Makhrov A.A., Verspoor E., Artamonova V.S., O'Sullivan M. 2005. Atlantic salmon colonization of the Russian Arctic coast: pioneers from the North America // Journal of Fish Biology V. 67 (supplement A). P. 68-79.
Parrish D.L., Behnke R.J., Gephard S.R., McCormick S.D., Reeves G.H. 1998. Why aren't there more Atlantic salmon (Salmo salar)? // Canadian Journal of Fisheries and Aquatic Sciences V. 55 (supplement 1). P. 281-287.
Rousset F. 1997. Genetic differentiation and estimation of gene flow from F-statistics under isolation by distance // Genetics V.145. P. 1219-1228.
Saisa M., Koljonen M.-L., Gross R., Nilsson J., Tahtinen J., Koskiniemi J., Vasemagi A. 2005. Population genetic structure and postglacial colonization of Atlantic salmon (Salmo salar) in the Baltic Sea area based on microsatellite DNA variation // Canadian Journal of Fisheries and Aquatic Sciences V. 62. P. 1887-1904.
Slatkin M. 1993. Isolation by distance in equilibrium and non-equilibrium populations // Evolution V. 47.
P. 264-279.
Svendsen J.I., Alexanderson H., Astakhov V.I., Demidov I., Dowdeswell J.A., Funder S., Gataullin V., Henriksen M., Hjort C., Houmark-Nielsen M., Hubberten H.W., Ingolfsson O., Jakobsson M., Kjsr K.H., Larsen E., Lokrantz H., Lunkka J.P., Lysa A., Mangerud J., Matiouchkov A., Murray A., Moller P., Niessen F., Nikolskaya O., Polyak L., Saarnisto M., Siegert C., Siegert M.J., Spielhagen R.F., Stein R. 2004. Late Quaternary ice sheet history of northern Eurasia // Quaternary Science Reviews V. 23. P. 1229-1271.
Tonteri A., Titov S., Veselov A., Zubchenko A., Koskinen M.T., Lesbarreres D., Kaluzhin S., Bakhmet I., Lumme J., Primmer C.R. 2005. Phylogeography of anadromous and non-anadromous Atlantic salmon (Salmo salar) from northern Europe // Annales Zoologici Fennici V. 42. P. 1-22.
GENETIC STRUCTURE OF ATLANTIC SALMON (SALMO SALAR) POPULATION OF THE WHITE SEA BASIN REVEALED WITH MICROSATELLITE MARKERS
A.K. Tonteri1, A.Je. Veselov2, A.V. Zubchenko3, J.I. Lumme4, S.M. Kalyuzhin5, M.Yu. Ozerov1, C.R. Primmer1
1Department of Biology, Division of Genetics and Physiology, University of Turku, Finland 2Institute of Biology, Karelian research Centre, Petrozavodsk, Russia 3Knipovich Polar Research Institute of Marine Fisheries and Oceanography, Murmansk, Russia 4Department of Biology, University of Oulu, Finland 5Varzuga Research Centre of Polar Ecosystems, Varzuga, Russia e-mail: mikhail.ozerov@utu.fi
Atlantic salmon (Salmo salar) populations spawning in the rivers of White sea basin are among the last European populations which are not majorly affected by human activities. Thus they form an important source of biodiversity for the entire European region in addition of being of immense economic and social importance to the local people. Here, microsatellite analysis of an extensive set of Atlantic salmon populations from the White and Barents Sea area was carried out in order to gain an insight into the population genetic structure based on nuclear markers. The populations were separated into four distinct clusters and individual assignment accuracy at the regional level (Atlantic and West Barents Sea; Kola Peninsula; Western White Sea; Eastern Barents Sea) was relatively high (87-96% success). These four groups can be considered as a good starting point for defining four distinct management units in the region, each of which may require differing management strategies as distinct patterns of genetic diversity and divergence characteristics were detected in different groups. Furthermore, the data obtained here can be used to differentiate individuals caught in offshore fisheries on a regional level with a relatively high degree of accuracy.
Еще по теме ГЕНЕТИЧЕСКАЯ СТРУКТУРА ПОПУЛЯЦИИ АТЛАНТИЧЕСКОГО ЛОСОСЯ (SALMO SALAR) БАССЕЙНА БЕЛОГО МОРЯ, ВЫЯВЛЕННАЯ С ПОМОЩЬЮ МИКРОСАТЕЛЛИТНЫХ МАРКЕРОВ[2]:
- ЛИПИДНЫЙ СОСТАВ ТКАНЕЙ АТЛАНТИЧЕСКОГО ЛОСОСЯ SALMO SALAR L. КОЛЬСКОГО ПОЛУОСТРОВА
- НЕРЕСТОВО-ВЫРОСТНОЙ ФОНД И СОСТОЯНИЕ ВОСПРОИЗВОДСТВА ЛОСОСЯ (SALMO SALAR) В МАЛЫХ РЕКАХ КУТОВОЙ ЧАСТИ КАНДАЛАКШСКОГО ЗАЛИВА
- СРАВНЕНИЕ ЛИПИДНЫХ СПЕКТРОВ СЕГОЛЕТОК АТЛАНТИЧЕСКОГО ЛОСОСЯ Salmo salar L. ИЗ ДВУХ БИОТОПОВ ПРИТОКА АРЕНЬГА (р. Варзуга, Кольский п-ов)
- ИЗУЧЕНИЕ ВОЗДЕЙСТВИЯ КОМПОНЕНТОВ БУРОВЫХ РАСТВОРОВ НА АКТИВНОСТЬ ФЕРМЕНТОВ СЕГОЛЕТОК СЕМГИ (SALMO SALAR L.)
- О ЛЕТНЕМ ИХТИОПЛАНКТОНЕ ДВИНСКОГО ЗАЛИВА БЕЛОГО МОРЯ
- 11. Нефтегазоносный бассейн Северного моря
- О ГЕНЕТИЧЕСКОМ И МОРФОЛОГИЧЕСКОМ РАЗНООБРАЗИИ MYTILUS EDULIS КАНДАЛАКШСКОГО ЗАЛИВА БЕЛОГО МОРЯ
- ВЛИЯНИЕ РАЗНОЙ СОЛЕНОСТИ НА ЛИПИДЫ АМФИПОД БЕЛОГО МОРЯ
- СОСТАВ КРИОФЛОРЫ ПРИБРЕЖНЫХ ЛЬДОВ КАНДАЛАКШСКОГО ЗАЛИВА БЕЛОГО МОРЯ
- СОСТОЯНИЕ ГИДРОБИОЦЕНОЗОВ РЕК ПОМОРСКОГО ПОБЕРЕЖЬЯ БЕЛОГО МОРЯ
- СЕЗОННО-ОБУСЛОВЛЕННЫЕ ИЗМЕНЕНИЯ В ТРАНСМИССИИ ПАРАЗИТОВ В ПРИБРЕЖЬЕ БЕЛОГО МОРЯ
- КРИОФАУНА ЛЬДОВ ПРОЛИВА ВЕЛИКАЯ САЛМА КАНДАЛАКШСКОГО ЗАЛИВА БЕЛОГО МОРЯ
- СТРУКТУРА И ДИНАМИКА ПРИМОРСКИХ РАСТИТЕЛЬНЫХ СООБЩЕСТВ ПОМОРСКОГО И КАРЕЛЬСКОГО БЕРЕГОВ БЕЛОГО МОРЯ Л.А. Сергиенко
- ДИНАМИКА СТРУКТУРЫ ПОСЕЛЕНИЯ MYTILUS EDULIS НА ЛИТОРАЛИ КАНДАЛАКШСКОГО ЗАЛИВА БЕЛОГО МОРЯ
- МОРФОЛОГИЧЕСКИЕ АСПЕКТЫ ВИДООБРАЗОВАНИЯ И ФОРМИРОВАНИЯ АДАПТАЦИОННЫХ СТРАТЕГИЙ ВИДОВ Р. COCHLEARIA L. В ПРИБРЕЖНЫХ РАСТИТЕЛЬНЫХ СООБЩЕСТВАХ БЕЛОГО МОРЯ Л.А. Сергиенко
- ПОЛОВАЯ СТРУКТУРА МОРСКОЙ ЗВЕЗДЫ ASTERIAS RUBENS L. НА ТЕРРИТОРИИ КАНДАЛАКШСКОГО ЗАЛИВА БЕЛОГО МОРЯ В 2003 ГОДУ
- Южные моря — сферы напряжённости: союзы в бассейне Тихого океана
- РАЗМЕРНЫЙ СОСТАВ И ЛИПИДНЫЕ ПОКАЗАТЕЛИ У АМФИПОД БЕЛОГО МОРЯ В РАЗНЫХ УСЛОВИЯХ ОБИТАНИЯ Г.А. Шкляревич1, В.В. Богдан2
- ПРОДУКЦИОННЫЕ ХАРАКТЕРИСТИКИ ФИТОПЛАНКТОНА БЕЛОГО МОРЯ В ЗАВИСИМОСТИ ОТ ИСТОЧНИКА АЗОТА