15.1. Анатомо-функциональная организация зрительных путей у детей в различные возрастные периоды
Топическая диагностика церебральных зрительных поражений у детей раннего возраста невозможна без знания анатомических и физиологических особенностей зрительных путей в различные возрастные периоды, а также принципов нейроофтальмологической диагностики поражений пре- и постгеникулярных путей у взрослых.
Экстраполяция стандартных ситуаций, встречающихся в практике офтальмологов, работающих с пациентами старшего возраста и применяющих широкий диапазон психофизических тестов, нередко позволяет установить правильный диагноз маленькому ребенку с церебральными зрительными поражениями, у которого использование традиционных психофизических исследований невозможно [Mackie R.T., McCulloch D., 1995; Haussler М. et al., 19961. Знание симптоматики поражений задних зрительных путей у взрослых помогает оценить функциональные проблемы, возникающие у детей раннего возраста с аналогичными патологическими состояниями. Некоторые поведенческие отклонения, развивающиеся у детей с церебральными зрительными поражениями, можно объяснить различными дефектами центрального и/или периферического зрения. Например, эксцентричное положение головы по отношению к рассматриваемому объекту может быть обусловлено гемианопти- ческими дефектами поля зрения [Jan J.E., 1991; Рогго G. et al., 1998].Зрительные пути простираются от сетчатки до зрительных центров затылочной коры. Прегеникулярный отрезок зрительных путей образуется ГКС (третий нейрон зрительного пути) и их аксонами, формирующими зрительные нервы, хиазму и зрительные тракты. Ход аксонов в зрительном нерве строится по принципу ретинотопично- сти: волокна, идущие от периферии сетчатки, располагаются по краям поперечника нерва, папилломакулярный пучок — в нижненаружном квадранте орбитального отдела до выхода из нерва центральных сосудов, а затем занимает центральное положение, сохраняя его в интраканаликулярном и краниальном отделах [Трон Е.Ж., 1968].
Орбитальный отдел зрительного нерва, составляющий у новорожденных в среднем 17 мм, удлиняется у взрослых ЛО 35 мм, интраканаликулярный — с 3 до 5 мм, а интракраниальный — с 4 до 6 мм. Одновременно происходит утолщение зрительного нерва: площадь поперечного сечения интракраниальной части составляет у новорожденного в среднем 4,27 мм2, в период с 3 до 4 лет — 8,9 мм2, у взрослых — 9,93 мм2 [Бирючков Н.В., 1963; Rimmer $. et al.,№93].
На основании мозга зрительные нервы образуют хиазму, где происходит частичный перекрест аксонов. Перекрещиваются аксоны, идущие от ганглиозных клеток, расположенных в носовых отделах сетчатки. Перекрещенный, филогенетически более старый пучок волокон определяется в хиазме уже к моменту рождения. Неперекрещенный пучок менее выражен и приобретает характерную волокнистость к 1—1,5 годам жизни [Троп Е.Ж., 1968].
У задней поверхности хиазмы > ^р' ■ начало зрительные тракты, заканчивающиеся в наружных коленчатых телах (НКТ) и подушках зрительных бугров. Длина зрительных трактов у новорожденных составляет около 15 мм. Их рост в длину происходит до
18— 25 лет, когда они достигают 30— 40 мм. НКТ — основной подкорковый центр зрительного анализатора — подразделяется на вентральную и дорсальную части. В вентральной части НКТ расположены два крупноклеточных (магноцеллюлярных) слоя, в которых образуют синапсы аксоны крупного диаметра, идущие от ганглиозных клеток сетчатки (ГКС) большого размера (М-клетки). В дорсальной части находятся четыре мелкоклеточных (парво- целлюлярных) слоя, в которых заканчиваются тонкие аксоны, идущие от мелких ГКС (Р-клетки) [Atkinson J., 1992]. Протяженность НКТ к моменту рождения составляет примерно 6,5 мм, у взрослых - 7-8 мм [Трон Е.Ж., 1968].
Обнаружены пути от НКТ к задней лимбической области, верхнему двухолмию, подушке зрительного бугра, претектальной зоне, заднему ядру зрительного бугра, а также к зрительному тракту и сетчатке. Не исключается существование перекрещенного пути через мозолистое тело в противоположное полушарие [ Школьник- Яр-
рос Е.Г., 1971].
Таким образом, в рамках сенсорной зрительной системы действуют два основных параллельно функционирующих проводника информации (рис. 15.1). Первый из них — тонковолоконные медленно проводящие парвоцел- люлярные, или П-клеточные, пути, начинающиеся от мелких ГКС и передающие информацию по тонким аксонам к синапсам в парвоцеллюлярных слоях НКТ. Затем эти волокна идут через зрительную лучистость к синапсам в слое 4с области Ѵ| и в конечном итоге проходят вентромедиально к нижней височной коре (область Ѵ4). Этот путь имеет значение для обработки информации, относящейся к точному пространственному разрешению, цвету, различению размеров, высококачественному стереозрению и, следовательно, к распознаванию объектов [Livingstone M., 1990]. Поражения данной подсистемы зрительного пути, включая область V4, приводят к прозо- пагнозии и церебральной ахроматопсии |Wa!l M., 1996].
Второй путь — крупноволоконная быстро проводящая магноцеллюляр- ная или М-клеточная система аксоны ГКС которой достигают синапсов магноцеллюлярного слоя НКТ. Затем
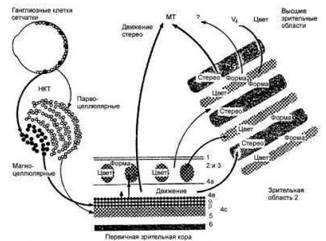
Рис. 15.1. Функциональная сегрегации и зрительной системе приматов [Livingstone М., 1994]. Объяснение в тексте.
волокна идут через зрительную лучистость к синапсам слоя 4с затылочной коры (Ѵ|), впоследствии — через корковую область Ѵ3 и, проходя дорсо- латерально в височной доле, достигают области МТ (Ѵ5). Этот путь определяет восприятие движения и мелькания в области высоких временных частот [Livingstone M., 1990]. С патологией этой системы ассоциируется синдром Balint [Wall М., 1996].
Парвоцеллюлярный путь объединяет около 80 % аксонов зрительного нерва, магноцеллюлярный — приблизительно 10 %. Оставшиеся 10 % аксонов имеют проекции в структурах среднего мозга [Plant G.T., Hess R.F., 1987]. При поражениях определенной локализации может преимущественно повреждаться Р- или М-путь, и поэтому даже выраженные зрительные дисфункции могут не сопровождаться снижением остроты зрения [Лес- селл С, Ван Дален Дж.Т.У., 1983; Бер- тулис А.В., Глезер В.Д., 1990; Later- za A., Nappo А., 1987; Wolf J.E.
et al.,1996] .
Зрительный нерв формируется приблизительно на 7-й неделе гестации из развивающихся ГКС, а уже на 8-й неделе (18—22-я постсомитные стадии) его волокна достигают головного мозга [Cook C.S. et al., 1995]. Макроскопически хиазма дифференцируется уже на
19- й постсомитной стадии развития зародыша, зрительные нервы и тракты — на 22-й (54—55-е сутки), верхние бугорки четверохолмия — на 23-й [Савельев СВ., 2002].
В первые месяцы жизни происходит морфологическое созревание на всех уровнях зрительного анализатора: дифференциация макулы, миелиниза- ция зрительных путей, увеличение к 4-месячному возрасту количества ши- пиков на дендритах и соме нейронов, расширение зрительных полей коры и формирование корковых петров, усиленный синаптогенез в зрительной коре больших полушарий головного мозга, в результате которого к 8-му месяцу жизни достигается максимальная синаптическая плотность [Авдеева Н.Н. идр., 1993; ФарберДА. идр., 1990; GareyL.I., 1984|. Перечисленные изменения приводят к быстрому увеличению остроты зрения у детей грудного возраста, что находит свое отражение в увеличении амплитуды, укорочении латентности и формировании компонентного состава ЗВП [3 нс лина Н.Н., 1987; ФильчиковаЛ.И., 1999].
Миелинизация зрительного нерва начинается приблизительно на 32-й неделе гестации в области хиазмы от дистального конца аксона к проксимальному [Cook C.S. et al., 1995]. Орбитальная порция зрительного нерва миелинизирована уже к моменту рождения. Быстрая миелинизация зрительных путей происходит до 2 лет жизни, позднее ее темпы значительно снижаются [Magoon R.H., Robb R.M., 1981].
В диске зрительного нерва, который располагается в склероальном канале, выделяют 3 части: внутренний слой нервных волокон, преламинарную и ламинарную части. При офтальмоскопии диск зрительного нерва у маленьких выглядит бледным. Диаметр диска у новорожденного составляет примерно 0,8 мм, у взрослого — 1,2— 1,9 мм. Вертикальный диаметр диска превышает горизонтальный в среднем на 0,18 мм.
В центре диска в области физиологической экскавации, которая у здоровых детей первых 3—5 лет жизни, как правило, не выражена, в глаз входит центральная артерия и выходит центральная вена сетчатки [Аветисов Э.С. и др., 1987]. Диск состоит из аксонов, астроцитов и соединительной ткани. Конвергенция аксонов ГКС от периферии к диску проявляется увеличением толщины слоя нервных волокон сетчатки в перипапиллярной зоне.При офтальмоскопии в бескрасном свете слой нервных волокон виден благодаря ярко выраженной исчерченно- сти, обусловленной ре флектированием света. Наибольшей толщины слой нервных волокон достигает в перипапиллярной зоне на 6 и 12 часах — в среднем 320 мкм: примерно 115 мкм приходится па астроциты и капилляры, а около 205 мкм — на аксоны. В носовой половине сетчатки и в зоне, лежащей между фовеа и диском, толщина слоя нервных волокон значительно меньше - 40-60 мкм [Quigley НА. et al., 1982]. При патологии (глаукома, атрофия или неврит зрительного нерва) исчезает характерная исчерченность слоя нервных волокон сетчатки, что связано с потерей аксонов и уменьшением его толщины на 50—75 % [Mun- tean Р., 1991; Radius R.L., Anderson D.R., 1979; Sharpe J.A., Sanders M.D., 1975; Tytia M.E., BuncicJ.R., 1990].
Зрительный нерв содержит около 1 млн аксонов различного калибра, группирующихся приблизительно в 400 отдельных пучков. Преобладают аксоны малого калибра, составляющие 90 % от общего количества. Аксонов большого диаметра значительно меньше — примерно 10 %. Аксоны, различающиеся по диаметру, идут от морфологически различающихся друг ОТ друга групп ГКС. В норме за 1 год теряется около 5000 аксонов, причем с возрастом происходит преимущественная потеря аксонов малого диаметра [Mikelberg F.S. et al., 1991]; возможно, благодаря этим процессам увеличивается и диаметр физиологической экскавации.
В раннем возрасте депривация, обусловленная заболеваниями зрительной системы приводит к изменению размеров клеток НКТ, причем клеточные нарушения более выражены при монокулярной депривации, чем при бинокулярной [Guillery R.W., 1973].
У приматов выделяют 2 сенситивных периода, отличающихся реакциями зрительных путей на депривацию. Первый продолжается с момента рождения до 8 нед жизни. Основные взаимодейст-между зрительными путями каждого из глаз в этом периоде — конкуренция и сегрегация корковых колонок глазодоминантности. Монокулярная депривация в этом возрасте приводит первоначально к гипертрофии не- депривиро ван його НКТ, а позднее — к параллельному сморщиванию как де- привированных, так и недепривиро- ванных парвоцеллюлярных клеток. Устранение депривации в эксперименте приводит к их частичному восстановлению (!), что имеет значение при определении оптимальных сроков реабилитации. Второй сенситивный период начинается с 8-й недели жизни и продолжается до 12—18 мес. В этой фазе для нормального формирования зрительного анализатора необходимы кооперативные взаимодействия между зрительными путями, идущими от каждого из глаз. При отсутствии таких взаимодействий происходит селективное уменьшение одного или обоих НКТ (депривированного и недеприви- рованного) [Sloper J.J., 1993].
T.A.Nealey и J.H.Maunsell (1994) изучали влияние избирательной инактивации магно- или парвоцеллюлярных отделов НКТ на активность нейронов стриарной коры путем регистрации зрительных ответов от поверхностных слоев V|. Инактивация парвоцеллюлярного отдела приводила к снижению нейрональной активности в поверхностных слоях Ѵ[, но эффект магноцеллюлярной блокады был более выраженным. Были также обнаружены нейроны, принимающие сигналы от обеих подсистем. Авторы отметили, что сигналы от маг- ноцеллюлярных путей привносят больший вклад в ответы, регистрируемые от поверхностных корковых слоев. Эти результаты — аргумент против упрошенного схематического картирования субкортикальных магно- и парвоцеллюлярных путей как дорсального и вентрального потоков обработки зрительной информации в коре.
Локальные поражения области V; (известной также как стриарная кора,
17- е поле, первичная зрительная или калькаринная кора) приводят к развитию скотом, для которых типичны гомон им ность и «конгруэнтность». Эти дефекты располагаются как бы противоположно по отношению к зоне поражения V|, так как хрусталик переворачивает изображение, попадающее на сетчатку, а назальные волокна зрительного нерва в последующем перекрещиваются в хиазме. Поражение макулярного представительства в области V, приводит к формированию центральной или «близко расположенной» (вследствие смешения точки фиксации) парацентральной скотомы. Такие дефекты приводят к нарушениям фиксации, функции сканирования и способности восприятия деталей.
Поражения V, ниже калькаринной щели приводят к развитию верхней квадрантопсии, что связано с инверсией изображения хрусталиком. Наоборот, поражения Ѵ|, локализующиеся выше калькаринной щели, обусловливают развитие нижней квадрантопсии [Wall М., 1996]. У пациентов с гомо- нимной гемианопсией может сохраняться «монокулярное височное полу- луние» — периферический височный сектор с одной стороны. Его сохранность в некоторых случаях может объяснять наличие так называемого слепого зрения, а также феномен Ridoch, при котором пациенты способны различать движение периферическим зрением слепого полуполя.
Изолированные поражения области V, у детей практически не встречаются. При поражениях, сопровождающихся изменениями области Ѵ| и белого вещества мозга в прилегающих областях и экстрастриарных зонах, отмечаются полиморфные комбинированные зрительные расстройства. Экстрастриарные дисфункции характеризуются менее грубой симптоматикой. Исключением из этого правила являются поражения 18-го и 19-го полей (по Бродману), при которых имеют место квадрантоптические (Ѵ|-подобные) дефекты в поле зрения даже в случае сохраности области Ѵ| и зрительной лучистости [Horton J.C., Hoyt W.F., 1991].
Повреждения затылочной коры могут вызывать нарушения стереозрения. Y. Takayama и М. Sugishita (1994) у 3 здоровых волонтеров исследовали нарушения стереозрения, индуцированные транскраниальной магнитной стимуляцией, используя красно-зеленые очки и случайные точечные стереограммы. Стимуляцию осуществляли при помощи магнитной катушки, расположенной по средней линии на
3— 4 см выше inion в проекции затылочных долей тангенциально поверхности черепа. У всех испытуемых обнаружены нарушения стереоскопического восприятия в период стимуляции. Параметры воздействия, вызывающего нарушения стереозрения, — стимулы длительностью не менее 0,2 с и частотой 10 Гц и выше. Авторы сделали вывод о том, что верхние зоны коры затылочных долей играют важную роль в стереозрении.
Некоторые слепые пациенты способны распознавать признаки стимулов, которые не соответствуют информативным возможностям их зрительных функций. Данное явление описано в литературе под названием «слепое зрение». Одни авторы предполагают, что перцепция в таких случаях опосредуется ретинотекгальной зрительной системой [Stoevig Р., Cowey А., 1989; Celesia G. et al., 1991], другие считают слепое зрение проявлением резидуальных функций крошечных функционально сохранных островков в пределах геникуло-калькаринных путей [Fendrich R. et al., 1992; KerkhoffG. et al., 1994].
Новый метод для оценки больных со слепым зрением был предложен R.Fendrich и соавт. (1992), использовавшими аппарат регистрации движений глзз при помощи фигур Пуркинье в комбинации со стабилизатором изображения, обеспечивающим точную проекцию стимула на сетчатку, для исследования поля зрения по стратегии предпочтительного взора у больных с левосторонней гомонимной гемианопсией и сохранением макулы. Тестирование позволяло выявить «спрятавшиеся» в гемианоптическом дефекте поля зрения островки остаточного восприятия, не обнаруживаемые при стандартной автоматической периметрии. Стимулы, предъявляемые в этих островках, могли быть различимы, хотя субъект утверждал, что не видит их. Наличие островков подразумевает сохранность корреспондирующих участков функционирующей коры в пределах пораженной области. Эти данные ставят под сомнение факт наличия «экстрастриарного» восприятия у больных со слепым зрением. P.J.Brent и соавт. (1994) подтвердили справедливость гипотезы R.Fendrich (1992) в сообщении о больном с резидуальным зрением, который отрицал возможность какого-либо цветовосприятия, но при тестировании методом предпочтительного взора идентифицировал цвета.
Согласно предложенной J.C. Horton и W.F. Ноу) (1991) карте, отражающей корковое представительство поля зрения, его центральная часть диаметром 10° занимает приблизительно 50 % площади стриарной коры. Их данные подтвердили R. McFadzean и соавт. (1994), также исследовавшие корреляцию результатов периметрии, КТ и М РТ у пациентов с поражениями стриарной коры.
Р. Krolak-Salmon и соавт. (2000) исследовали корреляцию между протяженностью резецированного у пациентов с эпилепсией участка головного мозга в рострокаудальном пространстве латеральной части височной доли и послеоперационными дефектами поля зрения. Размеры хирургического дефекта измеряли в ходе операции и позднее при помощи МРТ. Верхние гомонимные дефекты поля зрения, обусловленные повреждением зрительной лучистости, на контралатеральной резекции стороне выявлены у 83 % пациентов. Площадь дефектов поля зрения была прямо пропорциональна размерам резецированного участка. На остроту зрения операция не влияла. Результаты авторов подтвердили классические представления о топографии зрительной лучистости и ее ретинотопической организации. Строение зрительной лучистости объясняет некоторые периметрические феномены, вызывающие затруднения при их интерпретации в ходе нейроофтальмологического обследования.
Важно отметить, что дискомфорт, связанный с наличием дефектов в поле зрения, зависит от их локализации. Ни у одного из 15 пациентов с двусторонними верхними гомонимными дефектами поля зрения (различной степени тяжести — от минимальных до полной квад- рантопсии), обусловленными повреждением зрительной лучистости вследствие резекции височной доли, не было зрительного дискомфорта. Острота зрения у этих пациентов оставалась нормальной [Kroiak-Salmon Р. et al., 20001. Наоборот, при дефектах поля зрения, вовлекающих нижние квадранты, пациенты испытывают неудобства [Jacobson D. М., 1997]. J.D.Trobe и соавт. (1973), анализируя жалобы 104 пациентов с изолированной гомоним ной гемианопсией без сопутствующей офтальмо- и неврологической симптоматики, отметили, что 14 % из них не ощущали каких-либо зрительных нарушений.
J. /іГі! (1995) обнаружил корреляцию между локализацией гемианопти- ческих дефектов в поле зрения и дислексией. У пациентов с правосторонними парафовеальными дефектами способность к чтению была нарушена в большей степени, чем при левосторонних изменениях. У больных с левосторонними дефектами в поле зрения были нарушены преимущественно возвратные движения глаз к началу читаемой строки, тогда как при наличии правосторонних дефектов отмечались пролонгирование времени фиксации, уменьшение амплитуды саккадических перемещений вправо и регрессия сак- кад. Анализ продолжительности фиксации, амплитуды саккад глаз вправо и их обоюдной зависимости выявил уменьшение пространственных размеров перцептуального окна (интервала чтения) и значительное увеличение его временного объема [ZihlJ., 1995].
S.M. King и соавт. і исследовали способность к определению, локализации и различению высококонтрастных вспыхивающих стимулов и мелькающих или двигающихся объектов у 5 пациентов после гемисферэктомии. Для контроля за влиянием светорассеяния интенсивность порогов для монокулярно предъявляемых объектов в гемианоптическом поле была сопоставима с порогом, достигаемым при использовании дополнительных окк- людоров, перекрывающих слепую половину сетчатки тестируемого глаза. По аналогичной программе был обследован больной с односторонней деструкцией стриарной коры. В фотопиче- ских условиях никто из пациентов с гемисферэктомией не смог определить изменения освещенности в нарушенной части поля зрения, в то время как больной с монолатеральной деструкцией стриарной коры мог определять, различать и локализовать стимулы, а также оценить перемещение стимула через среднюю линию. При уменьшении фоновой освещенности пороговая интенсивность стимула, необходимая пациентам, перенесшим гемисферэктомию, для его монокулярного определения была идентична той, которая требовалась им для определения тест-объектов в условиях, когда вся зрительная информация была перекрыта окклюдором для слепого поля и могла быть доступна для зрительной системы лишь благодаря эффекту светорассеяния. Наоборот, у дестрииро- ванного больного монокулярный порог чувствительности в слепой части его поля зрения был значительно ниже, чем при предъявлении стимулов, в тот момент, когда дефектная часть поля перекрывалась окклюдором. Так как не было обнаружено каких-либо стимулов, которые пациенты после гемисферэктомии могли бы определять, а также не было выявлено взаимосвязи с различиями, обусловленными светорассеянием, авторы сделали вывод о том, что субкортикальные зрительные пути, уцелевшие после гемисферэктомии, не могли опосредовать -л: :о' к- поведенческие ответы по отношению к зрительной информации, возникающей в ге- мианоптическом поле [King S.M. et al., 1996].
Определенные дефекты в поле зрения могут потенциировать у пациентов глазодвигательные нарушения. Например, у больных с центральными дефектами в поле зрения вследствие нейро- офтильмалогичсскои іштолоіміі отмечено появление инверсивного оптокинетического нистагма. У пациентов с инверсивным оптокинетическим нистагмом, который провоцируется при помощи стимулов с более низкой пространственной частотой, в поле зрения выявляют центральные дефекты, имеющие большую площадь ]Сгс- vils L., van Vliet A.G.M., 1991].
Таким образом, зрительная система у детей претерпевает значительные анатомические и функциональные изменения в первые годы жизни В связи с этим характер и степень тяжести зрительных расстройств у детей с церебральными зрительными поражениями зависят не только от локализации и рас пространенности патол огичес кого процесса, но и от возраста, в котором развилось заболевание.
15.2.
Еще по теме 15.1. Анатомо-функциональная организация зрительных путей у детей в различные возрастные периоды:
- Заболевания задних зрительных путей у детей: терминология, диагностические критерии, классификация
- Острый неврит зрительного нерва и зрительных путей (код Н46)
- Основные закономерности возрастных изменений анатомо-оптических элементов глаза
- Вопрос № 22. Зрительный гнозис и его мозговая организация. Зрительные агнозии.
- I л а в a 1 5 ПОРАЖЕНИЯ ПОСТГЕНИКУЛЯРНЫХ ЗРИТЕЛЬНЫХ ПУТЕЙ У ДЕТЕЙ РАННЕГО ВОЗРАСТА ИМ. Молин
- Вопрос № 23. Зрительно-пространственный гнозис и его мозговая организация. Зрительно-пространственные агнозии.
- Сенсорные нарушения работы зрительной системы при поражении периферического, подкорковых и коркового зееньев зрительного анализатора (снижение остроты зрения, различные формы гемианопсий, скотомы, расстройства светоощущения, цветоощущения, фотопсии).
- Эпидемиология церебральных зрительных поражений у детей
- Принципы электрофизиологической диагностики церебральных зрительных поражений у детей раннего возраста
- Особенности развития зрительного анализатора у недоношенных детей
- Задержка зрительного созревания у грудных детей, дифференциальная диагностика и тактика ведения
- Проблемы реабилитации детей с церебральными зрительными поражениями
- К проблемам контроля возрастной динамикипсихического развития детей
- Клинические и функциональные критерии для диагностики церебральных зрительных поражений у младенцев
- Состояние зрительного анализатора у детей с ретинопатией недоношенных
- Возрастная организация